
Sample Parietal Lobe Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
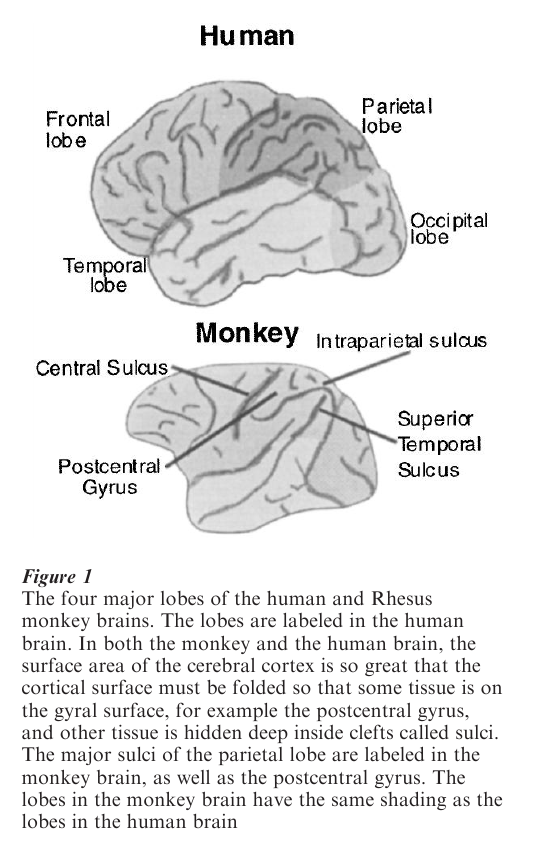
The parietal lobe is the part of the cerebral cortex lying between the occipital and frontal lobes, and above the temporal lobe (see Fig. 1). It is named for its proximity to the parietal bone, the wall of the skull (from the Latin paries, wall). It has five major functions: the primary analysis of somatic sensation (touch, the position of the limbs, temperature); the analysis of space using all sensory modalities; the specification of spatial targets for the motor system; the generation of attention; and the analysis of visual motion. Our first understanding of parietal function arose from the analysis of the deficits of patients with lesions of the parietal lobe (Critchley 1953). Subsequent insight into the mechanisms of parietal function came from neuroanatomical and neurophysiological studies.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
The importance of the parietal lobe was apparent to neurologists such as Hughlings Jackson, Dejerine, Head, and Holmes early in the twentieth century. They discovered that humans with parietal lesions had difficulty with tactile perception on the side of the body opposite the lesion (the contralateral side): they were not completely unable to feel stimuli, as they might be after a lesion of the spinal cord, but they had relative deficits: they might be able to feel touch, but not pain or temperature; they might perceive touch but have difficulty localizing the stimulus; they might have difficulty knowing the position of a limb in space; they might have difficulty identifying an object placed in their hand without looking at it—for example, they would not be able to reach into a pocket and pull out a key, leaving coins behind. Stimuli were often perceived to be less strong on the affected side than on the normal side of the body, and comparative discriminations were coarser. Sometimes patients who can appreciate a stimulus on an affected limb when it is the only stimulus touching them cannot appreciate it when two stimuli are applied simultaneously, one on the affected limb and the other on the normal limb. The most dramatic parietal deficit is anosognosia—patients, especially those with right parietal deficits, are entirely unaware of the limbs on the contralateral side: they think that a leg in bed with them belongs to someone else.
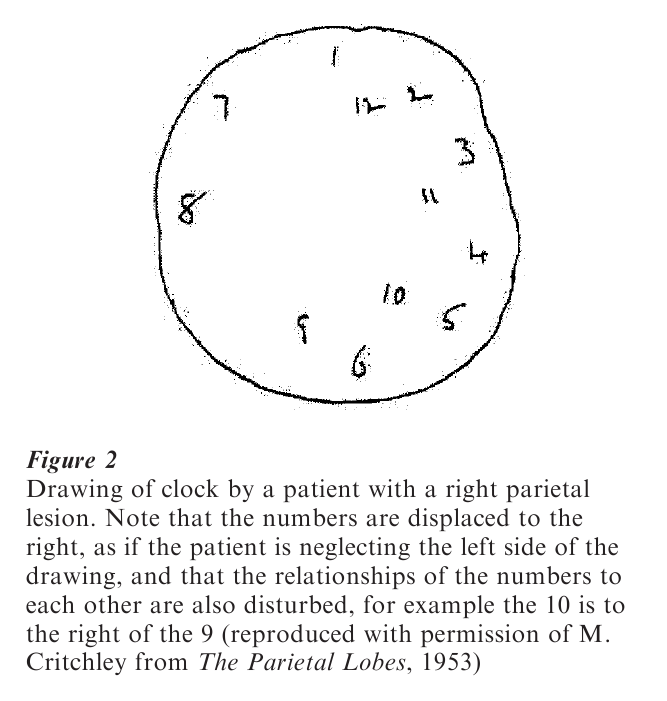
Patients with parietal lesions do not have deficits limited to the tactile or somatosensory domain: they have striking visual deficits as well. Patients with right parietal damage tend to neglect the left visual field, the left side of space relative to the midline of their bodies, and the left side of objects (see Fig. 2). When they draw a clock they cram most of the numbers on to the right side of the clock, neglecting the left; when they attempt to bisect a line they place the midline near the right end of the line rather than the middle. After this florid neglect has recovered, right parietal patients often still neglect a stimulus in the affected visual field when another one is presented simultaneously in the normal visual field. This deficit is interpreted as a deficit in visual attention: patients with parietal lesions are less efficient at shifting their attention into the affected field than into the normal field.
Patients with parietal lesions often exhibit optic ataxia, the inability to coordinate their movements using visual cues. They have difficulty knowing where objects are relative to their body. They misreach for objects, and make inaccurate eye movements toward them. They also have difficulty transmitting visual information to the motor system. When humans reach for an object, they open their hand to the approximate width of the object before their hand arrives at the object. Thus, people reaching for a can of soda will open their hands wide to accommodate the can; people reaching for a pencil will open theirs narrowly, just to accommodate the pencil. Patients with parietal lesions cannot do this, but reach for all objects with a stereotyped hand posture. The brain mechanisms that drive limb movement have no access to the detailed visual aspects of a target, even though they might know the spatial location of the target. A remarkable aspect of parietal deficits is that, although patients do not know where an object is, they have no difficulty telling what that object is—they know, for example, that the examiner is holding a wristwatch in front of them, but cannot find it to reach for it. They exhibit constructional apraxia—when asked to duplicate a block design they are unable to place the blocks in proper relationship to each other, but instead leave them in an unordered jumble even though they choose the proper blocks. Patients with lesions of the parietal cortex on both sides of the brain have difficulty shifting their attention away from the current object. The result of this deficit is that they only describe one in the world at a time—they may see a dog’s nose but not its ears, for example. They cannot use eye movements to explore the visual world. Patients with small lesions of the parietal cortex occasionally exhibit a specific deficit for the analysis of visual motion and the transmission of this information to the motor system. They have deficits both with the motor aspects of visual motion, for example, they cannot follow a moving object with their eyes; and they have difficulty with the perception of motion—knowing the speed of an object, or judging the predominant motion of a moving random dot pattern.
Space is not represented in a unitary fashion in the parietal cortex. Instead it seems to be organized in concentric spheres, or workspaces, that correspond to the range of action of certain motor systems. Patients with small lesions of the parietal cortex may have these different workspaces affected differentially. Thus a patient who can localize a nearby stimulus by reaching out and touching it with an arm may not be able to localize a stimulus further away in the same direction by touching it with the beam of a laser pointer. These are not merely deficits in movement—the patients show perceptual distortion, such as an inability to bisect a line, in the affected workspace.
Recent studies in neuroanatomy and neurophysiology, especially those done with the Rhesus monkey, have been able to illuminate the brain mechanisms whose destruction causes the dramatic clinical symptoms of the parietal lobe. The brain is a network of information-processing cells called neurons. Each neuron serves as a node in this network, receiving information from other neurons, often in different areas of the brain, and transmitting this information to other neurons. By studying the cytology of the parietal lobe and its connections with other parts of the brain, neuroanatomists have been able to divide it into a number of functional subdivisions. Neurons transmit information by emitting a series of electrical impulses called action potentials. If a neuron responds to a stimulus it issues a burst of impulses in response to the appearance of the stimulus. By monitoring the electrical activity of neurons in the parietal lobe of monkeys that are awake and performing behavioral tasks, neurophysiologists have begun to understand the signal processing that enables the parietal lobe to perform its function in the generation of human behavior.
Physiological and anatomical studies have subdivided the parietal lobe into a number of distinct areas which perform the various different functions whose deficits appear in patients with parietal lesions. The most anterior part of the parietal lobe is the primary cortical area for somatic sensation. Most of the posterior parietal cortex, starting with the posterolateral bank of the intraparietal sulcus is concerned with space and attention. The most posterior parietal area, in the superior temporal sulcus, is important in the analysis of visual motion.
The primary somatosensory cortex consists of four subdivisions, cytoarchitectonic areas 3a and 3b, located in the posterior bank of the central sulcus, and areas 1 and 2 located on the surface of the postcentral gyrus, the brain surface between the central and intraparietal sulci. Each of these subdivisions receives projections from the somatosensory parts of the thalamus, the way station between the spinal cord and brainstem, and the cortex. Areas 3a and 2 are responsible for deep bone and muscle sensation, including the muscle spindles that measure muscle length, and joint sensors that describe limb position. Areas 3b and 1 are responsible for sensation from the skin. These areas project backward to area 5, on the postcentral gyrus and the anterior bank of the intraparietal sulcus. This area integrates the various primary sensory signals, and transmits somatosensory signals to the premotor cortex so the information can be used for not only the perception of somatic sensation but also the coordination of movement. Single neurons in the anterior parietal cortex describe the properties of tactile stimuli in a very precise manner: for example, Mountcastle showed that neurons responsive to vibration describe the intensity and frequency of that vibration so accurately that a computer could construct the nature of the stimulus by decoding the neural pulse train.
The posterior parietal cortex is subdivided into a number of areas. Those located in the intraparietal sulcus associate somatic sensation, vision, and audition for the analysis of space, and the description of this world for action. Many of the areas in the intraparietal sulcus describe particular workspaces (Colby and Goldberg 1999). The physiological properties of these areas have been studied extensively in the Rhesus monkey.
Perhaps the best-studied area of the monkey parietal cortex is the lateral intraparietal area (LIP). This area describes distant space, the space that primates explore with their eyes. It projects to the frontal eye fields and the intermediate layers of the superior colliculus, brain areas known to be important in the generation of the rapid eye movements (saccades) that human and nonhuman primates use to explore their environment. It also projects to areas in the temporal lobe, TE and TEO in the Von Bonin and Bailey nomenclature, which are important in the analysis of pattern. An important connection between the generation of saccadic eye movements and the analysis of the visual world is the process of attention, the selection of objects in the world for special analysis. In general, human and nonhuman primates pay attention to the targets for their eye movements, and saccades and attention are closely linked, but they can be dissociated in certain cases—for example juvenile Rhesus monkeys certainly pay attention to the alpha male of their troop, but they never look directly at him. Animals attend to stimuli either because the stimulus is relevant to the current task, or because the stimulus is naturally attention worthy, stimuli that are, in the words of William James, ‘big things, bright things, moving things, or … blood’ (James 1890). For example, stimuli that appear abruptly in the visual field inevitably draw visual attention.
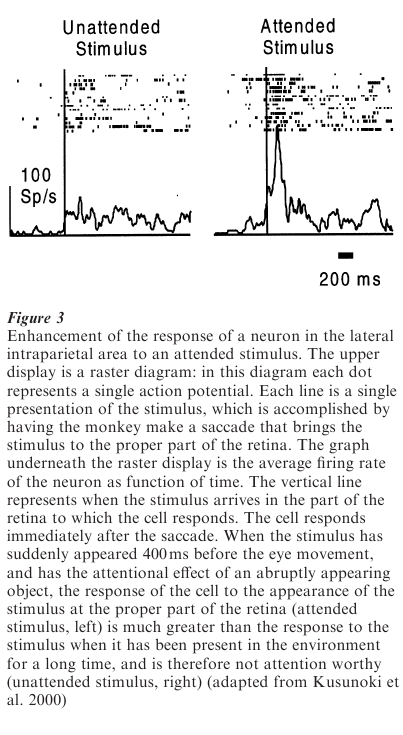
Neurons in LIP respond both to saccade targets and to attended stimuli that are not necessarily the targets for saccades. These neurons respond to visual stimuli, and their activity is modulated by whether the animal is paying attention to the stimulus. Neurons in LIP respond more to attended than to nonattended stimuli, independently of whether those stimuli are attended to because of their importance to the task or because they are in the class of stimuli that inevitably draw attention. Presumably it is the absence of this attentional modulation system that impairs the ability of parietal patients to attend to stimuli in the world contralateral to the lesion. Neurons in LIP also respond to sounds when the monkey attends to them (Anderson et al. 1997). A classic task in which to study the activity of neurons is the delayed saccade task.
In this task a monkey looks at a spot of light and another one flashes briefly in the visual field. A few seconds later the original fixation point disappears, and the monkey makes a saccade to the spatial location of the now vanished saccade target. Neurons in LIP respond to the stimulus, and then continue responding even though the stimulus has disappeared. This activity has been interpreted as either maintaining the memory of the saccade target, or focusing attention at the saccade target. It is probable that this area of the brain supports both functions: focusing attention on spatial locations, and providing the oculomotor system with information about the location of potential saccade targets. LIP serves as a model for the other, less well studied areas of the parietal lobe: neurons specify possible targets for the action appropriate for the workspace, as well as responding to attended objects in that workspace.
The most posterior part of the parietal cortex of the monkey lies in the superior temporal sulcus. This area is important for the analysis of visual motion. Neurons in the middle temporal area and the middle superior temporal area are specialized for the analysis of visual motion, and respond selectively to stimuli moving with certain speeds and direction. Electrical stimulation of these areas can affect monkeys’ perception of motion, and lesions in these areas render monkeys unable to follow moving stimuli with their eyes, or to make judgments about the quality of visual motion.
Bibliography:
- Andersen R A, Snyder L H, Bradley D C, Xing J 1997 Multimodel representation of space in the posterior parietal cortex and its use in planning movements. Annual Review of Neuroscience 20: 303–30
- Colby C L, Goldberg M E 1999 Space and attention in parietal cortex. Annual Review of Neuroscience 2: 319–49
- Critchley M 1953 The Parietal Lobes. Hafner, New York
- James W 1890 The Principles of Psychology. Holt, New York
- Kusunoki M, Gottlieb J, Goldberg M E 2000 The lateral intraparietal area as a salience map: the representation of abrupt onset, stimulus notion and task relevance. Vision Research 40: 1459–68
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality