
Sample Event-Related Potentials Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
The term event-related potential (ERP) refers to changes in the brain’s electrical activity locked in time to an externally defined event. ERPs have been used since the early 1960s, when they were usually referred to as ‘evoked potentials,’ to investigate sensorimotor and cognitive function in healthy and clinical populations. This research paper outlines how ERPs are acquired, what is known about the origin of ERP signals in the brain, and the sorts of question to which the ERP method can be applied. A comprehensive introduction to these and related issues can be found in Rugg and Coles (1995).
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. How Are ERPs Obtained?
ERPs are small in relation to features of the brain’s electrical activity (the EEG) that are not time-locked to a specific event. Therefore it is necessary to employ processing techniques that can extract the ERP ‘signal’ from the ‘noise’ in which it is embedded. The most commonly employed technique is signal averaging, whereby a number of EEG segments, each time-locked to the same class of event, are averaged to give a waveform in which consistent features of the segments (the ERP) are retained while features that vary across segments are attenuated (see Fig. 1). Depending on the relative sizes of the ERP and the background activity (the ‘signal-to-noise ratio’), the number of segments required for averaging might vary from as few as 10 to more than 1,000. It is important to note that the validity of the signal averaging procedure rests on the assumption that the ERP does not vary across segments to any significant extent. If this assumption is violated (e.g., because of habituation over successive segments or just poor time-locking to the eliciting event), the averaged waveform will not be representative of the time-locked activity elicited in the single segments from which it is derived.
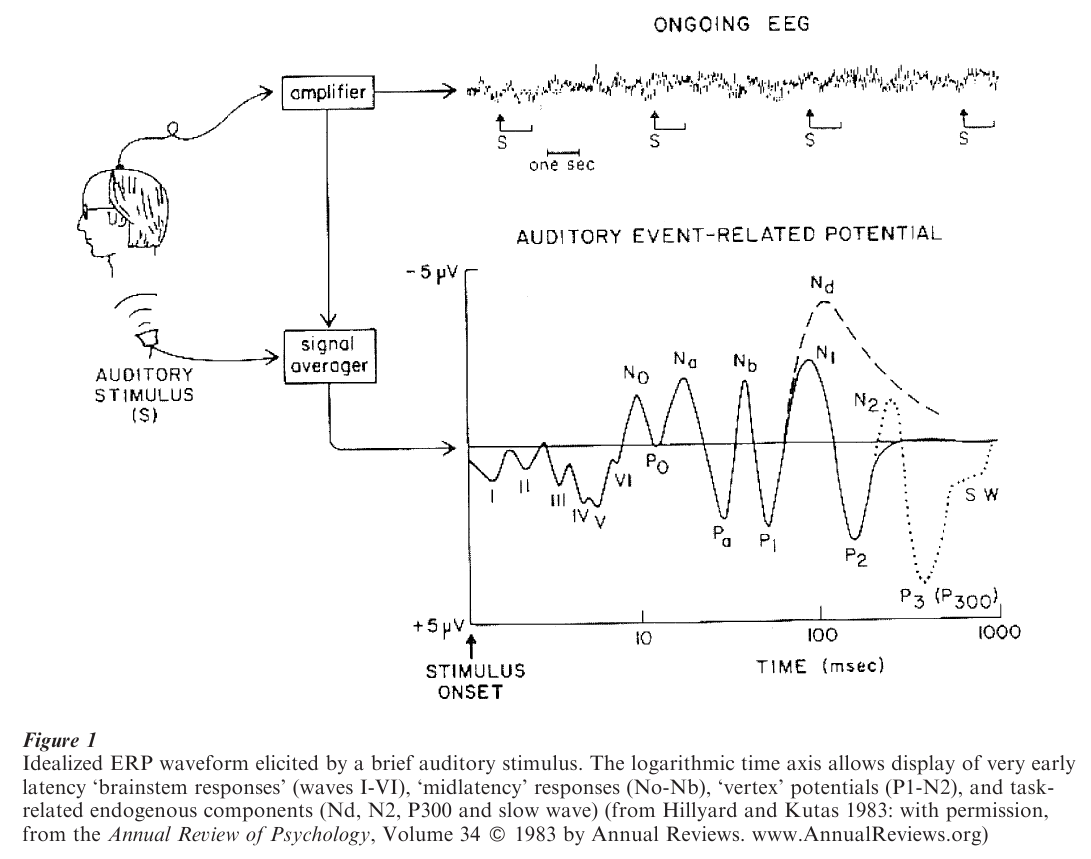
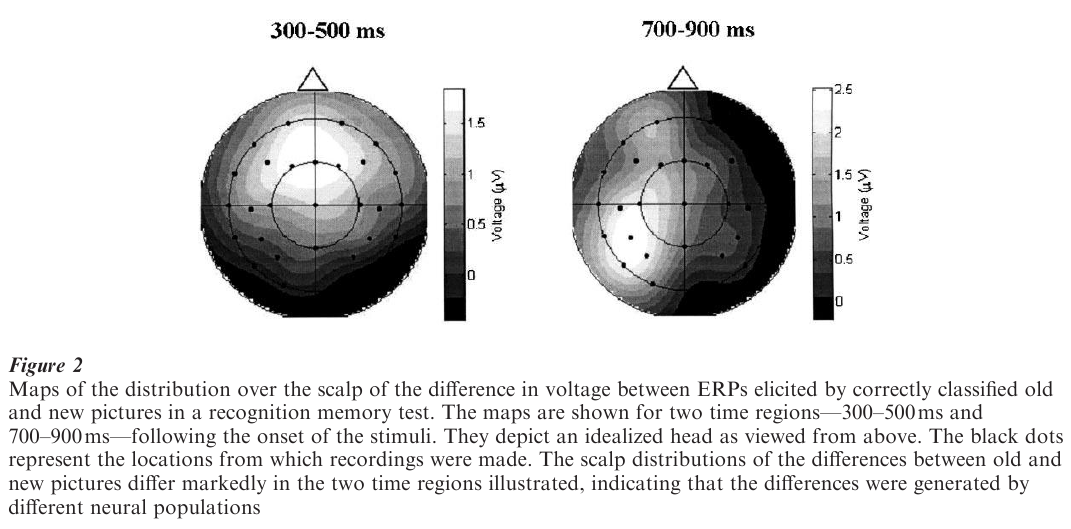
As is shown in Fig. 1, the ERP is conventionally depicted as a plot of voltage against time, so that the ERP waveform represents fluctuations over time in the potential difference between a recording site on the scalp and some ‘reference’ site such as the earlobe or nosetip. These fluctuations represent the time-varying changes in neural activity that were triggered by the eliciting event. The depiction of ERPs in this fashion emphasizes the good time resolution that can be achieved with the method, and in the days when intrumentation limitations meant that ERPs could be recorded from only a few sites at a time was sufficient to allow appreciation of the data. Technological advances mean that it is now commonplace to record from 100 or more electrodes simultaneously. The rich spatial information provided by such ‘dense-array’ recordings can best be appreciated by depicting data in the form of scalp ‘maps’ rather than as time-varying waveforms (see Fig. 2).
2. Generation And Source Localization
Because the electrical currents generated by single neurons are miniscule, scalp electrical fields must arise from neural populations in which the activity of the individual neurons is sufficiently synchronous to allow their currents to summate and be detectable at the scalp. It is generally believed that, under most circumstances, the duration of neuronal action potentials (‘spikes’) is too short to allow the currents associated with these potentials to summate in this manner. It is assumed therefore that most scalp electrical activity is generated by postsynaptic potentials, the duration of which is sufficient to allow summation even if their onset in different neurons is not perfectly synchronous.
Crucially, if activity in a neuronal population is to be detectable at the scalp, it must not only overlap in time, but also have certain spatial characteristics. Specifically, the neurons comprising the population must be arranged geometrically in a way that allows the currents in each neuron to flow in the same direction, so that the currents add to rather than cancel each other. Such a configuration is known as an ‘open field’ and generally involves the alignment of neurons in parallel. A good example of this sort of arrangement is given by the pyramidal cells of the neocortex, which are oriented at right angles to the cortical surface. Neurons in random or circular arrangements give rise to ‘closed’ fields, in which the currents generated by each neuron tend to cancel each other. Because the neurons in many brain structures have closed field configurations (for example, the basal ganglia), ERPs can never provide more than a partial record of the neural activity triggered by the event to which they are time-locked.
Although the principles underlying the generation of event-related scalp electrical fields are understood quite well, the localization within the brain of the regions responsible for a given field (the field’s ‘sources’) is a formidably difficult problem. The difficulty arises because the shape of a field on the scalp does not predict the configuration of its underlying sources in any simple way, and there is no mathematical method for uncovering this relationship unambiguously (indeed, from a mathematical perspective the problem is ‘ill-posed,’ i.e., it does not have a unique solution). Because of this, in all but a handful of cases it has not been possible to localize the generators of ERP phenomena with any great accuracy. Unlike researchers using other neuroimaging methods, ERP researchers have therefore tended to describe their data not in terms of the activity or function of specific brain regions or systems, but with respect to features of the ERP waveforms themselves and their presumed psychological correlates.
3. ERP Components And Their Classification
Although it is most natural to think of ERPs as representing brain activity which follows a given event (event-following potentials), ERPs can also be recorded during intervals preceding an event (event preceding potentials). An example of event-preceding potentials are those that precede a motor act (‘motor’ or ‘readiness’ potentials), providing valuable information about movement preparation and execution.
As can be seen from Fig. 1, an ERP waveform following a sensory event consists of a series of peaks and troughs, or ‘deflections.’ Early in the waveform, the timing (‘latency’) of these deflections shows relatively little variability across different individuals, and within an individual the size (‘amplitude’) and latency of the deflections are determined largely by the physical characteristics of the stimulus (characteristics such as intensity, duration, etc.). These early deflections—sometimes referred to as ‘exogenous’ potentials—reflect initial sensory processing of the eliciting event, and they are employed routinely in the clinical assessment of sensory function (Chiappa 1997). For example, subtle damage to the optic nerve can result in delay in the latency of exogenous visual ERPs.
As time since stimulus onset progresses, features of the ERP waveform are determined less by physical features of the stimulus and more by the psychological state of the subject and the nature of the processing accorded the stimulus. Indeed, from around 250–300 ms or so post-stimulus onset, psychological factors largely predominate, such that the same physical stimulus will elicit markedly different waveforms depending on such factors as its relative probability of occurrence, whether it signals the need to make a response, etc. Emphasizing their dependence on internal, psychological factors rather than external ones, late-onset ERP deflections are often referred to as ‘endogenous’ potentials.
The deflections observed in ERP waveforms are easy to measure and provide convenient descriptive labels; for example, ‘P1’ and ‘N2’ refer respectively to the first positive and second negative deflection in a waveform; ‘N250’ refers to a negative deflection peaking around 250 ms post-stimulus onset, etc. It is not the case, however, that each deflection represents the time course of a single source of neural activity within the brain. Because activity recorded at the scalp represents the sum of all the sources active at a given time, the peak of an ERP deflection need not reflect the moment when any one source is maximally active. For this reason a distinction is made between ERP deflections—the surface features of waveforms—and ERP components—contributions to the waveform arising from specific sources or source configurations, the spatial and temporal overlap of which yields the recorded waveform. An important part of ERP analysis involves the decomposition of recorded wave-forms into their constituent components.
4. Applications Of ERPs
As already mentioned, exogenous ERPs are widely used in the assessment of sensory function. Efforts have also been made to use endogenous components to assess cognitive function and its impairment in various clinical groups, albeit with only limited success as aids to clinical diagnosis. From the perspective of psychology and related disciplines, the ERP method is of interest largely to the extent that it can be used to further understanding of cognitive functions and their neural underpinnings. ERP findings can be informative in these respects for several reasons, only the most important of which are outlined here (see Rugg and Coles 1995). First, neural activity associated with stimulus processing can be measured with a temporal resolution in the millisecond range. Thus, the maximum time required by the nervous system to discriminate between different classes of stimulus (e.g., novel vs. familiar faces) can be determined directly, even when volunteers are not required to discriminate between or respond to the stimuli. Second, ERP waveforms can be formed ‘off-line,’ after experimental trials have been sorted into different conditions according to the subject’s behavior. It is therefore easy to compare records of brain activity associated with different classes of response (e.g., remembered vs. forgotten items in a memory test). Finally, ERPs can be used to investigate whether different experimental conditions engage distinct cognitive operations. On the assumption that different patterns of neural activity imply the engagement of different cognitive operations, it follows that when two ERP scalp distributions differ (as in Fig. 2), it can be concluded that the experimental conditions associated with each distribution engaged at least partially separate cognitive operations. Thus, differences in the spatial characteristics of ERP fields can be used to make inferences at a psychological level, even if the location of the neural activity responsible for these differences is unknown.
These applications of the ERP method are illustrated below with respect to two central areas of psychological enquiry. For further information on these and other areas of psychology and cognitive neuroscience in which ERPs have made a contribution see Kutas and Dale (1997) and Rugg and Coles (1995).
5. Selective Attention
The question of whether human selective attention acts ‘early’ or ‘late’ during stimulus processing has proved difficult to resolve on the basis of behavioral evidence alone. ERP data clearly show however that when processing load is high, selective attention exerts an effect early on during processing. This conclusion derives from the finding that, in all sensory modalities investigated, attended stimuli elicit exogenous ERP components that are larger in amplitude than the components elicited by unattended stimuli. In the auditory modality, for example, attention has been reported to influence components occurring as early as 20 ms post-stimulus onset, which reflect the earliest cortical stages of auditory processing (Woldorff and Hillyard 1991). Such findings have been interpreted as evidence that attention acts in part by selectively biasing stimulus processing before full perceptual analysis has occurred (Luck et al. 2000).
6. Language
Whereas the distinction between syntax and semantics is fundamental from a formal linguistic perspective, the question whether semantic and syntactic processes are distinct in terms of their underlying cognitive operations is the subject of debate. In 1981 it was shown that the terminal words of semantically ‘anomalous’ sentences (e.g., ‘he took his coffee with cream and socks’) elicited a negative-going ERP deflection (the ‘N400’ component) that was much enhanced relative to ERPs elicited by nonanomalous endings (e.g., ‘it was his first day at work’; Kutas and Hillyard 1980). Subsequent work showed that this effect was present in the auditory modality also, and could be found for words at any position within a sentence. It was also shown that the amplitude of the N400 component depended on a word’s semantic expectancy, rather than whether or not it was anomalous (e.g. check elicits a larger N400 than stamp following ‘he mailed the letter without a check stamp’). Later, it was demonstrated that syntactic anomalies (e.g., ‘the broker persuaded to sell the stock’ vs. ‘the broker hoped to sell the stock’) also elicit characteristic ERP deflections, but with quite different distributions over the scalp to the N400 and, for one such deflection (the ‘P600’ component or ‘syntactic positive shift’), a difference in polarity (Osterhout and Holcomb 1992). These deflections also do not depend on anomaly per se; was in the ‘garden-path’ sentence ‘the lawyer charged the defendant was lying’ elicits an enhanced P600 relative to the same word in ‘the lawyer charged that the defendant was lying,’ presumably reflecting the need for the listener or reader to engage in ‘syntactic repair’ at this point in the sentence. These and related findings have been taken as evidence in support of the idea that the derivation of syntactic and semantic information during sentence comprehension depend on distinct neural and cognitive processes (Hagoort and Brown 1999).
7. Relation To Other Methods
The ERP method is only one of several methods permitting the noninvasive measurement of human brain activity. In particular, the method of functional magnetic resonance imaging (fMRI) has recently come to prominence as a powerful way to identify event related neural correlates of sensorimotor and cognitive function (D’Esposito et al. 1999). Whereas fMRI has far superior spatial resolution to that possible with ERPs, the temporal resolution of fMRI is markedly inferior. Since an understanding of brain function requires that neural activity be located accurately in time as well as in space, ERPs are likely to continue to play an important role in cognitive neuroscience for some time to come. Indeed, the development of methods for the integration of ERP and fMRI data would represent a major step forward for functional imaging (McCarthy 1999).
Bibliography:
- Chiappa K H 1997 Evoked Potentials in Clinical Medicine, 3rd edn. Lippincott Williams & Wilkins, Philadelphia, PA
- D’Esposito M, Zarahn E, Aguirre G K 1999 Event-related functional MRI: Implications for cognitive psychology. Psychological Bulletin 125: 155–64
- Hagoort P, Brown C M 1999 The neurocognition of syntactic processing. In: Brown C M, Hagoort P (eds.) The Neurocognition of Language. Oxford University Press, Oxford, UK, pp. 271–316
- Hillyard S A, Kutas M 1983 Electrophysiology of cognitive processing. Annual Review of Psychology 34: 33–61
- Kutas M, Dale A 1997 Electrical and magnetic readings of mental functions. In: Rugg M D (ed.) Cognitive Neuroscience. Psychology Press, Hove, UK, pp. 169–242
- Kutas M, Hillyard S A 1980 Reading senseless sentences: Brain potentials reflect semantic incongruity. Science 207: 203–5
- Luck S J, Woodman G F, Vogel E K 2000 Event-related potential studies of attention. Trends in Cognitive Sciences 4: 432–40
- McCarthy G 1999 Event-related potentials and functional MRI: A comparison of localization in sensory, perceptual and cognitive tasks. Electroencephalography and Clinical Neurophysiology Suppl. 49: 3–12
- Osterhout L, Holcomb P 1992 Event-related potentials elicited by syntactic anomaly. Journal of Memory and Language 31: 785–806
- Rugg M D, Coles M G H (eds.) 1995 Electrophysiology of Mind: Event-Related Brain Potentials and Cognition. Oxford University Press, Oxford, UK
- Woldorff M G, Hillyard S A 1991 Modulation of early auditory processing during selective listening to rapidly presented tones. Electroencephalography and Clinical Neurophysiology 79: 170–91
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality