
Sample Color Vision Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. iResearchNet offers academic assignment help for students all over the world: writing from scratch, editing, proofreading, problem solving, from essays to dissertations, from humanities to STEM. We offer full confidentiality, safe payment, originality, and money-back guarantee. Secure your academic success with our risk-free services.
Color has long interested people from many different professions: artists, engineers, physicists, psychologists, and biologists, all with their own viewpoints and areas of primary interest. We consider here how the underlying neural organization of the eye and brain allows us to discriminate color and produces our various color percepts.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
The light from every location in space varies in two physical dimensions: intensity (or the number of photons) and wavelength (or the energy levels of the photons). Color vision is the ability to discriminate changes in the spectral composition of a visual stimulus independently of its effective intensity, and vice versa. Because color vision apparently evolved independently several times, it is an ability that we share with many other species. However, we will restrict our discussion to the color vision of humans and other Old World primates.
1. Trichromatic Limitation
A visual scene may include millions of different local spectral distributions of light. A human observer with normal color vision, however, can visually match every one of these millions of different colored stimuli by appropriately combining and adjusting the intensities of just three different chromatic lights. The three variable-intensity lights that can be combined to match any color are known collectively as primaries. Despite what one may have been taught, there is no requirement that the primaries be red, blue, and yellow, or any other particular set, and they can be drawn from similar colors or from very different colors. The only restriction is that no two primaries can be combined to match the third. This trichromacy implies that at some processing stage, color information is limited by being transmitted through a three-channel pathway. It does not identify the point at which the three-channel bottleneck occurs. Many earlier theories assumed that trichromacy implies the existence of only three cone types, but recent evidence indicates that the limited number of different channels to the brain, not the number of receptors, limits our color vision to three dimensions. The trichromatic limitation to our color vision, the collapse of millions of physical dimensions into just three perceptual dimensions, should not be taken to mean that we are poor at discriminating colors. We make very fine discriminations within this three-dimensional space and can distinguish millions of different colors. However, the colors we see are not related to the physical spectral characteristics of objects in any simple fashion.
To gain any useful information from the retinal image is a much more difficult task than one might imagine; it requires multiple levels of neural computation involving billions of cells that occupy about a third of the whole cortex. An understanding of some aspects of this complex computational problem helps explain some otherwise-puzzling features of color vision.
2. Separating Objects From The Illuminant
One problem faced by the visual system is that most natural objects are visible not because they themselves emit light, but only indirectly because they reflect some proportion of whatever light falls on them. The light that reaches our eyes from a given object is, therefore, a product of the intensity and spectrum of the light source and the reflectance characteristics of the object’s surface. Since we do not have any independent information about the illuminant, it is not possible to determine the reflectance characteristics of any region, its color or its lightness, with certainty. The visual system can only guess at the solution. The main way it does so is by assuming that the illuminant is relatively constant whereas objects vary across space and time (with eye movements). It then estimates the intensity and spectral distribution of each region by comparing the activity evoked by the light from this region with the average neural activity (assumed to be primarily a function of the illuminant) over small neighboring retinal regions and across 100 msec or so of time. As a result, the perceived color of an object depends not only on the light coming from it but also on the light coming from nearby points in space, as well as the light to which the eye has been adapted recently. Among the consequences of this process are the well-known color and brightness contrast effects.
3. Separating Intensity And Wavelength Information
A second problem for the visual system is that a photoreceptor can only report how many photons it has absorbed, and the number of photons absorbed is determined both by the number of incident photons (the intensity of the light) and by their respective energy levels (the wavelength of the light). Thus, intensity and wavelength are completely confounded in the receptors. The information, for instance, that 100 photons were absorbed—the only thing that the receptor can report—does not allow one to determine either the intensity or the wavelength of the light in that region. To determine either the wavelength or the intensity requires neural comparisons of the outputs of neighboring receptors and of different cell types in the cortex.
4. Conflict Between Acuity And Color Vision
A third difficulty results from the fact that the visual system, over at least part of the retina, needs to maintain as high spatial acuity as possible. To build a detailed spatial map requires treating each receptor within the high-acuity area as a separate, equivalent spatial unit. However, it requires a comparison of the outputs of at least two receptors that differ in their spectral sensitivities at each location to separate intensity and wavelength information. There is then an inherent conflict between the requirements for color and those for high spatial acuity.
We do not yet fully understand how the visual system handles these and other problems, but we do understand some of the processes that contribute to the solutions in the early stages of visual processing. We will describe briefly the sequence of steps, as we understand them to date, in the neural processing in the retina and visual cortex that lead to our ability to identify and characterize the color and lightness of objects.
5. Photopigments And Spectral Sensitivity
The initial stage in the visual response is the absorption of light by the photopigment molecules in the rod and cone receptors. Variations in the amount of light absorbed by the photopigments in a receptor trigger a complex series of reactions, leading ultimately to synaptic transmission of the information to the next neurons in the pathway to the brain. Each receptor type is differentially sensitive to light of different wavelengths. The basic shapes of the spectral sensitivity functions of the various cone photopigments are similar, but the spectral region to which they are most sensitive differs among the S, M, and L (for short-, medium-, and long-wavelength) cones (see Fig. 1A). The cone photopigments all have quite broad spectral sensitivity. The L and M pigments in particular absorb significantly at all visible wavelengths. The wavelength of the light is thus not given by which receptor is activated, since all are activated by lights of most visible wavelengths, but only by the relative activation of the three different receptor types.
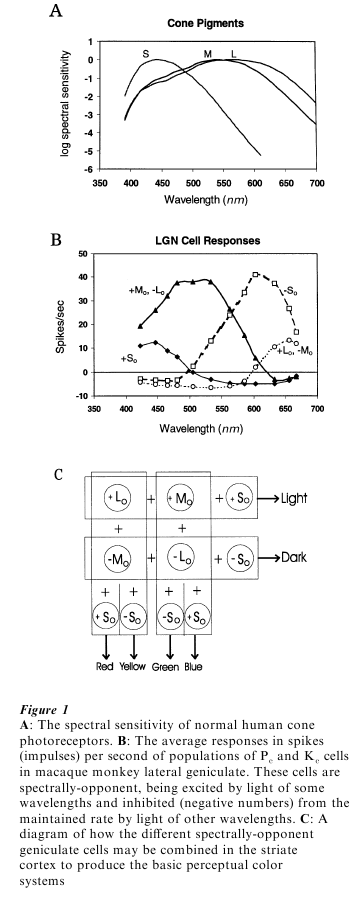
The compromise made by the visual system between the requirements for spatial vision (all cones the same) and for color vision (cones differing in spectral sensitivity), can be seen in the fact that the spectral sensitivities of the L and M cones are very similar whereas the S cone curve is very different. Almost all our spatial vision is based on the L and M cones, with the S cones contributing just to color vision.
6. Color Vision Deficits
Almost all human observers appear to have the same S-cone photopigment, but some differ in their L and M cone pigments. About 7 percent of human males and perhaps 1 percent of females have so-called red-green color vision deficiencies. The most extreme of these color-defective individuals (very few are truly color-blind, although that term is often used) are missing either the L or the M cone pigment and therefore have only two cones types. They are thus dichromats, and can match any color using only two primaries. Protanopes are missing the L cone pigment, and deuteranopes the M cone pigment. With only the S cones and either the L or M cones, these dichromats can distinguish short from long wavelengths, but they confuse colors over the whole red-yellow-green portion of the spectrum, where the S cones are insensitive.
More common than dichromats are anomalous trichromats, in whom either the L or the M cone pigment is present but shifted in spectral sensitivity with respect to that in normal observers. They thus are trichromats, but they make different color matches than normals do. Finally, there are very rare individuals, tritanopes, who are missing the S cones and are, therefore, dichromats.
7. Genetic Basis Of Color Vision Deficits
The prevalence of red–green color-vision defects and their higher incidence in males can be accounted for by the chromosomal location of the various photo-pigment genes. The S cone pigment gene is on chromosome 7, and tritanopia occurs equally frequently in both sexes. The M and the L cone photopigment genes, however, lie end-to-end on the X chromosome. Because they are very similar and are in adjacent positions on the same chromosome, the L and M photopigment genes are subject to inter-changing parts. This results in a variety of L and M genes in the population. The extreme case is one in which the two genes end up being the same, thus specifying only an M or an L photopigment. Since males only inherit one copy of the X chromosome while females inherit two, a female would have to inherit anomalous genes from both parents to be color-defective. A male, on the other hand, only receives a single X chromosome, from his mother, and is correspondingly more likely to have anomalous or dichromatic color vision. An important corollary of this is that many females appear to have more than just two LM pigments, and thus more than three different cone photopigment types. Nonetheless, females with more than three cone types still have only a trichromatic color vision system. Presumably the two L cone types that such a woman might possess are treated as being identical by later neural processes and thus do not produce an additional chromatic dimension. This strongly suggests that the trichromatic limit to human color vision is the result of only three types of analyses in the retina, and of only three paths from there to the cortex, not, as was long supposed, because of the presence of only three cone types.
8. Receptor Function
The retina is part of the brain that just happens to lie some distance away, in the eye, and receptors are similar to neurons elsewhere in the nervous system. They respond to stimuli by either a depolarization or a hyperpolarization of their outer membranes, and they communicate by releasing synaptic chemicals in proportion to the amount of depolarization in the synaptic region. While ordinary neurons respond to external transmitter chemicals, however, photo-receptors respond to chemicals generated within the receptors themselves, triggered by the absorption of photons by their photopigments.
The photopigment molecules are in the outer segments of the photoreceptors. The receptor outer segment membrane is maintained at an intermediate level of polarization by a balance between opposing sets of chemical interactions. The effect of increases and decreases in the amount of light absorbed by the photopigment molecules (due to a change in either the intensity or the wavelength of incident light) is to shift the polarization balance briefly in one direction or the other. This polarization change in the outer segment is conducted down the receptor to its synaptic region where it produces a change in the rate of synaptic chemical release. A decrement in light absorbed shifts the balance towards depolarization and leads to an increase in synaptic transmitter release to activate the next neurons in the chain to the brain; an increment in light absorption shifts the balance towards hyper-polarization and produces a decrease in transmitter release. As we move our eyes around in a scene, the amount of light absorbed by each receptor oscillates up and down, and the receptors quite faithfully transduce these oscillations into oscillating amounts of synaptic transmitter release. Since these variations in absorption could be due to either intensity differences or wavelength differences in the scene, however, it is left to the neural processing to sort out which is which and thus identify the color and brightness of each visual object.
9. Retinal Connectivity
There are three main types of retinal processing and three main paths from the eye to the cortex, which we will refer to as the Mc, Pc, and Kc paths. They are so called because they pass through the magno-, parvo-, and konio-cellular layers of the lateral geniculate nucleus, the relay station between the eye and the cortex. None of the cells in these three paths signals the amount of light in its region of the retina. Rather, each cell signals the differences between different distributions of light within its receptive field (RF), the region from which it receives its input. The cells do not signal the outputs of just one cone type, either, but rather they signal the sums and differences between the activity of different cone types.
The cells in the Mc path, in a spatially-opponent organization, signal changes in the amount of light absorbed by a small group of L and M cones relative to the amount absorbed by the L and M cones in a larger, overlapping area. Recall that the L and M cones absorb across the whole visible spectrum. Half the cells in this path fire if there is more light in the RF center than in the surround (+LMc -LMs); the other half fire if there is less light in the center than the surround (+LMc -LMs). With uniform illumination, center and surround responses will tend to cancel. This organization begins to emphasize information from objects and minimize that related to the illuminant, because if all the L and M cones in a region absorb more light, or if they all absorb less, it is probably due to a change in the illumination (as when the sun goes behind a cloud or emerges from one). The reflections from different parts of an object, on the other hand, will likely differentially activate one group of cones relative to their neighbors. Information about such intensity variations is what is captured and carried to the cortex by cells in the Mc pathway.
The cells in the Kc path, in a spectrally-opponent organization, signal changes in the amount of light stimulating one or more S cones relative to the amount stimulating the neighboring L and M cones ( +Sc -LMs). We shall refer to these as S-opponent cells, or +So. Since the cone photopigments each have broad spectral sensitivity, one cannot identify the wavelength by which cone is activated. However, differences in the relative amount of activation of the different cone types is related to the spectral distribution of the light, and this information is captured by a spectrally-opponent organization. Short-wave-length light is absorbed relatively more by the S than by the L and M cones. It will therefore excite a cell in the Kc path. Long-wavelength light is absorbed more by the L and M cones than by the S cones, and will thus inhibit a +So cell in the Kc path.
Finally, cells in the Pc path (about 80 percent of the total) have both a spatially-and a spectrally-opponent organization. These cells, at least in the central retina, receive input from a single L or M cone in their RF center, and a small group of L and/or M cones in the surround. There are thus four varieties of these cells:+ Lc -Ms (or Lo), -Lo, +Mo, and +Mo. The +Lo and -Mo cells have the same spectral response characteristics (both L-M), as do the +Mo and -Lo cells, (both +M-L), and are therefore combined in Fig. 1B. Since they encode the difference between the activity of L vs. M cones in spectrally-opponent organizations, they respond to color differences in the long-wavelength half of the spectrum. (It is the absence of these cells, because of a lack of either L or M cones, that makes a protanope or deuteranope unable to discriminate red from yellow from green.) However, these cells also signal changes in activation of a single L or M cone relative to that in the surrounding cones, in a spatially-opponent organization. A +Lo cell, for instance, fires to a large red patch, but also to a small light spot on a dark background. The Pc cells thus carry both color and intensity information. The neural processing in the retina begins but does not complete the process of separating color and intensity information.
10. Cortical Processing
The three main retino-cortical paths project separately to the striate cortex (also known as the first visual cortical area, V1), but within the cortex the paths are no longer discrete. Rather, V1 cells combine the outputs of the different cell types in various ways. Most of the early cortical processing involves building RFs to detect various spatial aspects of the pattern (e.g., spatial frequency, orientation, motion, depth) within a local cortical region. With respect to color vision per se, the primary processing involves separating color and luminance information, and further separating changes due to the illuminant from those due to visual objects, by lateral interactions over large regions.
To separate luminance and color information, the outputs of Pc cells are combined in two different ways. When their outputs are summed in one way, the luminance components to their responses sum and the color components cancel. Summed in a different combination, the color components sum and the luminance components cancel. Consider a striate cortex cell that combines inputs from one or more + Lo and +Mo cells in a region. The cortical cell would respond to luminance variations but not to color variations, since the neurons providing its inputs both fire to luminance increments in the RF center and to decrements in the surround, but the color organizations of its inputs are opposite to each other (one being L-M and the other M-L). Combined with input from a So cell, this would produce a V1 cell that fires to white (light increments) and inhibits to black (light decrements) but does not respond to pure color variations. This is represented in the top row of Fig. 1C. However, a V1 cell receiving inputs from both + Lo and -Mo cells, or from both Mo and Lo cells (columns in Fig. 1C), would respond to color changes but not to luminance variations since their color responses would add, but their luminance RFs, which are opposite to each other, would cancel. This organization by itself would produce L-M color cells that would fire to so-called warm colors (red and yellow) and inhibit to cool colors (blue and green). M-L cells would fire to cool colors and inhibit to warm colors. As shown in Fig. 1C, the further addition of +So or – So cells can split these classes into separate red and yellow, and separate blue and green systems, respectively.
All of the primary visual information is passed through V1, but subsequent visual areas are partially specialized for the further analysis of various different functional aspects of vision. One later visual area (V4) is crucially involved with color perception. Individuals with localized V4 lesions can still discriminate objects on the basis of their color variations, but they report that the objects now appear to have no hue, as if viewed on a black-white television screen. There is also a report of one case with the reverse loss: a patient who could see colored but not black-white objects.
11. Color Appearance
The appearance of a color can be specified by values along just three perceptual dimensions known as hue, saturation and brightness. Hue refers to the characteristic described by such color names as red, yellow, green, and blue. Saturation refers to the extent to which the stimulus differs perceptually from a purely achromatic (i.e., white, gray, black) axis. The third dimension is brightness or lightness. That our perceptual space is three-dimensional reflects the basic trichromacy of vision.
A normal observer can describe the hue of any light (disregarding surface characteristics) by using one or more of only four color names (red, yellow, green, and blue). These so-called unique hues form two opponent pairs, red–green and blue–yellow. Red and green normally cannot be seen in the same place at the same time; if unique red and unique green lights are added in appropriate proportions, the colors cancel and one sees a neutral gray. Orange can be seen as a mixture of red and yellow, and purple as a mixture of red and blue, but there is no color seen as a red–green mixture (or as a blue–yellow mixture). This perceptual opponency is also reflected in color contrast. Red can induce the appearance of green into neighboring regions, and after staring at a red surface one sees a green after-image. The yellow–blue opponent pair produces similar effects. It was these perceptual characteristics of color that led Ewald Hering in the nineteenth century to propose that the various color systems were not independent but rather that color was processed in a spectrally opponent organization, an idea which has since been amply verified in the presence, discussed above, of spectrally-opponent cells in the path from receptors to the cortex.
Bibliography:
- De Valois R L, De Valois R L 1988 Spatial Vision. Oxford University Press, New York
- Hurvich L M 1981 Color Vision. Sinauer Press, Sunderland, MA
- Kaiser P K, Boynton R M 1996 Human Color Vision. Optical Society of America, Washington, DC
- Neitz J, Neitz M 1998 Molecular genetics and the biological basis of color vision. In: Backhaus W G S, Kliegl R, Werner J S (eds.) Color Vision. Walter de Gruyter, Berlin, pp. 101–19
- Spillmann L, Werner J S 1990 Visual Perception: The Neuro-physiological Foundations. Academic Press, New York
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality