
Sample Motor Cortex Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
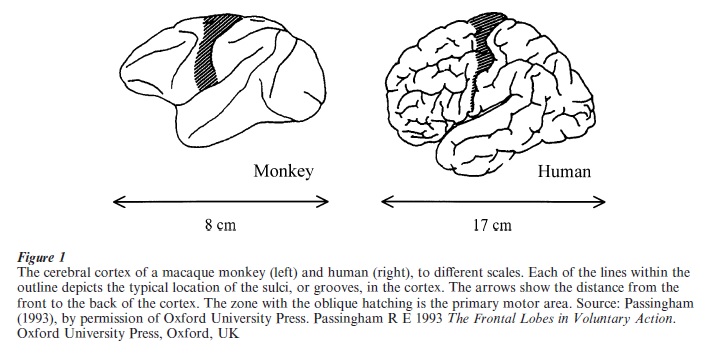
The term motor cortex has both a broad and a narrow sense. The broad meaning refers to a number of functionally distinct cortical areas, which collectively control voluntary action (Wise 1999). The narrow sense, adopted here, pertains exclusively to one of those areas, the primary motor cortex (Fig. 1). The primary motor cortex comprises only a small proportion of the cerebral cortex but plays a crucial role in controlling movement. Some of its neurons have axons that terminate directly on motor neurons in the spinal cord, giving motor cortex the most direct possible access to muscles (Porter and Lemon 1993).
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. History Of Motor Cortex
The motor cortex has a short history by evolutionary standards. Before the advent of mammals, about 200 million years ago, the motor cortex did not exist. Only mammals evolved a motor cortex in the sense used here. Accordingly, it probably functions to supplement the remainder of the vertebrate motor system, which evolved more than 500 million years ago.
In contrast to its relatively short evolutionary history, on a geological timescale, the motor cortex has a long intellectual history by the standards of neurobiology. In the fourth and fifth centuries BC, Hippocratic physicians observed that damage to one side of the brain caused motor disabilities on the opposite side of the body (Lassek 1954). In the nineteenth century, G. Theodor Fritsch and Eduard Hitzig in Germany and David Ferrier in England confirmed experimentally the existence of a localized motor cortex. They showed that electrical stimulation of the motor cortex in dogs and monkeys produced movements on the opposite side of the body. Although condemned by antivivisectionists of the time as inapplicable to humans, this late nineteenth-century animal research established the principles of cortical organization for all mammals, including our species.
2. Description Of Motor Cortex
Like other areas of the cerebral cortex, the motor cortex consists of several cell layers. However, the motor cortex exceeds other areas in thickness. Four other characteristics collectively identify the motor cortex: electrically stimulating it evokes movements more easily than for other cortical areas; it lacks a layer common to most cortical areas (layer 4); it receives relatively direct inputs from the basal ganglia and cerebellum; and it lies immediately in front of the somatosensory cortex. The neurons that send axons directly to the spinal cord all arise from layer 5, including some of the largest neurons in the cerebral cortex. Named after Vladimir Betz, these giant neurons can reach 60 µm in cell-body width in monkeys and 120 µm in people. Cells in the motor cortex send outputs to and receive inputs from somatosensory and premotor cortex. Subcortical targets include the basal ganglia, cerebellum, red nucleus, and other parts of the brain stem and spinal motor system. According to contemporary theory, individual cortical cells function as components of distributed neural networks incorporating a number of anatomically distinct structures, rather than as part of a localized ‘center’ performing a certain function.
2.1 Nature Of The Body Map
By noting the progression of epileptic seizures from one body part to another, the British neurologist J. Hughlings Jackson surmised that the motor cortex was arranged in a rough map of the body. Electrical brain stimulation confirmed that conclusion. Nearest the top of the brain (Fig. 1), the leg is represented. Representation, in this sense, implies that this part of motor cortex processes information about leg movements. The leg representation sends some of its axons to motor neurons that control leg muscles. Adjacent to the leg representation, in roughly the middle of the hatched zone in Fig. 1 (from top to bottom), lies an arm representation. The part of motor cortex that controls the face, head, tongue, and lips is situated yet further from the top of the brain. The finer organization of the motor cortex remains controversial and, although many detailed maps have been proposed, none have stood the test of time. In this respect, the organization of motor cortex contrasts with that of the somatosensory, visual, and auditory areas of the cerebral cortex, which have finer and more orderly maps of the receptors for touch, sight, and hearing, respectively.
3. Function Of Motor Cortex
The earliest knowledge about the function of motor cortex arose from observing the effects of damage to it or its main output pathway, the pyramidal tract. Unfortunately, the medical literature often lacks an accurate assessment of the exact site and extent of brain damage. It is clear, however, that a number of severe motor disabilities follow such damage, including weakness, difficulty in controlling the muscles individually, and loss of fine motor control. It is commonly held that the motor cortex functions mainly to permit motor ‘fractionation,’ i.e., the independent control of muscle groups that usually work in concert. In this sense, the motor cortex can be viewed as supplementing the subcortical motor system. According to this idea, motor fractionation adds an important degree of flexibility to the motor system. Although in many primates, including humans, independent movement of the fingers typifies fractionation, the ability to fractionate movements is by no means confined to the digits. Whatever the motor cortex does, it does for the body as a whole.
In the early 1960s, the American neurophysiologist Edward Evarts developed a method for recording the activity of individual neurons in the motor cortex of awake, behaving monkeys. He began by investigating a question first posed by Jackson in the nineteenth century. Does the motor cortex control muscles or movements? This seemingly innocuous question engaged researchers for many years. It was finally resolved by dividing the question into two components, which yielded contradictory answers. One question concerns the addressing of muscles by motor cortex. Do cells in motor cortex address individual muscles (the ‘muscle’ view) or several muscles, which may act together to produce a coherent action (the ‘movement’ view). Early brain-stimulation studies suggested single-muscle addressing. However, with more careful research the multiple-muscle movement hypothesis prevailed for the specific issue of addressing. The other question concerns the signal communicated to these muscle groups. Does motor cortex control the level and pattern of force generated by muscles (the ‘muscle’ view) or, alternatively, joint angles, limb position, and the direction of movement (the ‘movement’ view)? Neurophysiologists have found that most cells in the motor cortex control muscle activity rather than movement. Motor cortex emits a force signal, which usually causes movement, but may instead generate force without movement (e.g., when one pushes against an immovable object). A smaller number of cells control a combination of force and movement.
3.1 Population Coding
Individual neurons have their greatest activity for force in one direction, with systematically less activity as the direction of force diverges from that preferred direction. The cells are thus said to be tuned for direction. On the assumption that these cells generate force in their preferred direction, it is possible to estimate a collective force signal emanating from a neuronal population. This computation, termed the population vector, matches closely the direction of movement and force generated in a straight line, as well as both spiral and sinusoidal trajectories (Georgopoulos 1991).
3.2 Motor Cortex, Volition, And Awareness
Voluntary actions are those that are learned, attended, and based on a comparison among alternatives (Passingham 1993). Other kinds of action include reflexes and other innate behaviors. It seems likely that the motor cortex controls voluntary actions, which are typically based upon consciously perceived information. However, this contention implies neither that it lacks a role in reflexes nor that all voluntary movements depend upon consciousness.
Recently, the distinction between explicit (or declarative, perceived) knowledge and implicit (or procedural, unperceived) knowledge has gained wide acceptance. The former has access to conscious awareness; the latter does not. Both explicit and implicit knowledge can guide voluntary action. For example, people with ‘blindsight’ can point to a visual stimulus while denying that they see it (Weiscrantz 1986). Similarly, one well-studied neurological patient can orient her hand in order to put it through a narrow slot, but cannot report the orientation of the same slot (Milner and Goodale 1995). These are examples of implicit, unperceived knowledge guiding voluntary action. It seems likely that the motor cortex is involved in both explicitly and implicitly guided voluntary movements.
It has become increasingly clear that the processing of sensory information for perception differs from that for movement. In some respects, the performance of the motor system surpasses the neural systems underlying perception. For example, people can make movements that accurately match the size of an object to be grasped, but report incorrectly the size of the same object due to a visual illusion (Milner and Goodale 1995). The motor system thus has access to somewhat different sensory information than the brain systems underlying perception. As a consequence, one cannot assume that people’s actions invariably reflect their perceived knowledge.
3.3 Motor Learning And Plasticity
Motor learning and memory include both adaptation and skill acquisition. Adaptations involve changes in motor performance without an enhancement in capabilities. For example, if a prism shifts the visual image of a target, it takes several attempts to correct the error induced in reaching toward that target. Such adaptation leaves the system able to reach or throw accurately, but no more so than before adaptation. By contrast, learning a skill involves obtaining a new capability or improved performance, beyond the system’s prior limits. Evidence from brain-imaging studies suggests that the motor cortex changes the way it processes information during both skill learning (e.g., learning a movement sequence) and adaptation (e.g., tracking a rotating target). Other methods lead to the same conclusion. During sequence learning, for example, the movements caused by stimulating motor cortex become larger and easier to evoke (Hallett 1999). And during motor adaptation, the activity levels and other properties of neurons change.
Nearly 1 percent of the population has suffered a stroke causing motor disability. Many of these patients survive for years, 35 percent for a decade or more. Recovery of function usually proceeds slowly and often remains incomplete. Recent research has shown that on account of the brain’s plasticity, important organizational changes occur after damage to motor cortex. Practice that leads to such plasticity enhances recovery of function (Nudo et al. 1997). Future research will likely focus on drug treatments and other therapies aimed at regulating cortical plasticity. The advent of treatments to ameliorate damage to the motor cortex could have effects resembling the development of correctional optics in the thirteenth century. Eyeglasses extended the productivity of artisans by freeing them from the limitations of middle-age farsightedness (Landes 1998). Extending the productivity of patients with motor cortex damage could have similar benefits for both the individual and society.
Bibliography:
- Georgopoulos A P 1991 Higher order motor control. Annual Review of Neuroscience 14: 361–77
- Hallett M 1999 Plasticity in the human motor system. Neuroscientist 5: 324–32
- Landes D S 1998 The Wealth and Poverty of Nations: Why Some are so Rich and Some are so Poor. 1st edn. W W Norton, New York
- Lassek A M 1954 The Pyramidal Tract: Its Status in Medicine. Thomas, Springfield, IL
- Milner A D, Goodale M A 1995 The Visual Brain in Action. Oxford University Press, Oxford, UK
- Nudo R J, Plautz E J, Milliken G W 1997 Adaptive plasticity in primate motor cortex as a consequence of behavioral experience and neuronal injury. Seminars in Neuroscience 9: 13–23
- Passingham R E 1993 The Frontal Lobes in Voluntary Action. Oxford University Press, Oxford, UK
- Porter R, Lemon R 1993 Corticospinal Function and Voluntary Movement. Clarendon Press, Oxford, UK
- Weiscrantz L 1986 Blindsight. A Case Study and Implications. Oxford University Press, Oxford, UK
- Wise S P 1999 Motor cortex. In: Adelman G, Smith B H (eds.) Encyclopedia of Neuroscience, 2nd edn. Elsevier, Amsterdam
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality