
Sample Neural Plasticity In Auditory Cortex Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
This research paper covers the topic of neural plasticity in the auditory cortex, particularly with reference to learning and memory in adult subjects. As used here, the term ‘neural plasticity’ refers to systematic long-term (minutes to months) changes in the responses of neurons to the same physical stimulus (e.g., a tone), due to experience.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
Neural plasticity in the auditory cortex is interesting not only in itself but also as a case study in the intersection of two scientific fields that had developed quite separately, those of sensory physiology and the neurobiology of learning and memory. Furthermore, this topic provides a clear example of how assumptions constrained thought and experiment for most of the twentieth century. Finally, because the majority of studies on learning and sensory systems have been carried out in the auditory cortex, the development of new ideas and findings in the auditory cortex has direct implications for studies of neural plasticity in adult somatosensory and visual cortices.
The history of neuronal plasticity in the auditory cortex involves four stages: (a) sensory-motor framework excluding the auditory cortex from learning and memory; (b) delineation of cortical responses to sounds in anesthetized animals; (c) revelation of plasticity of responses to sounds during learning; and (d) discovery that learning systematically changes the frequency tuning of neurons in the auditory cortex.
1. Traditional Functional Organization Of The Cerebral Cortex
Attempts to understand the auditory cortex (as well as many other brain systems and structures) have their origin in the nineteenth century, within the framework of a sensory-motor conception of the nervous system. In the 1820s, the French physiologist F. Magendie, building on the partially correct work of the English anatomist C. Bell, showed that the dorsal and ventral roots of spinal nerves have different functions, sensory and motor, respectively. In the 1840s, Marshall Hall, an English physiologist, discovered that the sensory and motor nerves, together with the internal substance of the cord, constituted a behavioral structural– functional unit, that of the spinal reflex. The question of whether or not the entire nervous system was organized on sensory-motor principles formed much of the research program for the rest of the century (Young 1970).
It was generally believed that the cerebral cortex was not electrically excitable and operated only in a holistic manner, as it was widely thought to be the location of the unitary self or soul. However, in 1870 Fritsch and Hitzig, two German medical students, used focal electrical stimulation to discover the motor cortex. Also during the last quarter of the nineteenth century, David Ferrier, an English physiologist, used brain lesions to approximately locate the auditory cortex in the temporal lobe, by showing impaired hearing in animals. Thus by the beginning of the twentieth century the sensory-motor principle had been extended to the cerebral cortex. However, many regions of the cortex appeared to be neither sensory nor motor. About 1910, several of these areas were labeled the ‘association cortex’ by the anatomist W. LeGros Clark. The dominant implication of this formulation was that sensory and motor processes converged in these areas to form learned associations. Implicit in this schema was the concept that sensory (and motor) regions of the cortex were not sites of learning and memory.
2. Auditory Cortical Responses To Acoustic Stimuli
The availability of sensitive electronic amplifiers in the 1930s enabled scientists to record brain potentials that were elicited by controlled sensory stimuli, hence initiating the modern field of sensory neurophysiology. In the United States, recordings in the 1940s by Woolsey and Walzl in the cat and by Tunturi in the dog delineated auditory cortical fields. In particular, their studies revealed that the primary auditory cortex exhibits a ‘tonotopic map,’ i.e., that cells best tuned to one frequency were located next to cells that were tuned to an adjacent frequency. In effect, the primary auditory cortex has an organization for acoustic frequency that is similar to that of a keyboard or a harp. Subsequent more detailed studies showed that single neurons exhibited tuning functions, responding best to one frequency (the ‘best frequency’) and less strongly to other, higher and lower adjacent frequencies. Tuning curves are also called ‘frequency receptive fields’ because they show the regions of acoustic frequency to which neurons are receptive, i.e., to which they respond (Fig. 1).
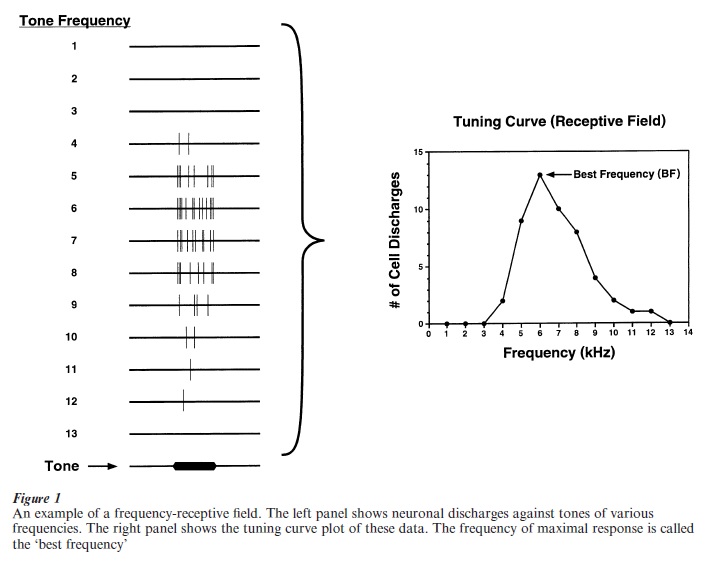
3. Initial Behavioral Studies Of Cortical Plasticity
While auditory neurophysiology was determining how the auditory cortex responds to sound, the field of learning and memory independently began to study how responses of the brain change when a stimulus becomes behaviorally important or loses its importance. Robert Galambos and his colleagues in Washington, DC performed a seminal experiment in 1955. Cats were classically conditioned by pairing an auditory conditioned stimulus (CS) with a puff of air to the face. Evoked potentials elicited by the CS became larger during conditioning; complementary studies of habituation, i.e., repeated presentation of a tone without reinforcement, by Irwin Westenberg, later revealed the opposite effect, a decrement of cortical responses to the repeated stimulus. Over the next decades, numerous additional studies in various animals and training situations demonstrated that learning systematically alters evoked potentials and neuronal discharges in the auditory cortex elicited by conditioned stimuli or habituated stimuli.
The findings in toto showed that the responses of the auditory cortex to sounds were affected not only by the physical sounds themselves, but also by the learned psychological or behavioral importance of acoustic stimuli. These findings clearly showed the error of the traditional belief that sensory cortices had purely sensory functions and were not regions directly involved in learning and memory.
However, the documentation of neural plasticity had little effect on the field of sensory neurophysiology, probably for several reasons. First, learning studies used only one or two tones, which were not interesting to sensory workers who used many stimuli. Second, there appears to have been conceptual confusion in which sensation was equated with perception so that the constructive aspect of the latter was not appreciated; hence plasticity of sensory responses was mistakenly seen as incompatible with perceptual accuracy. Neither was sensory cortical plasticity initially influential within the field of learning and memory itself. This relative neglect appeared to reflect the traditional belief in association cortex as sites of learning and memory on the one hand and emphasis on the hippocampus and other noncortical structures on the other hand.
4. Receptive Field Plasticity In Learning And Memory
The first evidence that learning actually changes receptive field (RF) tuning was obtained in 1986 at the University of California Irvine. As this initial experiment concerned so-called ‘associative’ areas of the auditory cortex, rather than the tonotopic auditory cortex, it did not address the question of whether learning actually changed tuning in the primary sensory cortex. Then in 1990 Bakin and Weinberger recorded RFs from cells in the primary auditory cortex before and immediately after conditioning to a tonal conditioned stimulus. They found that responses to the CS frequency increased whereas responses to the best frequency (BF), and many other frequencies, decreased. These opposing changes were sufficient to shift frequency tuning toward or to the frequency of the CS (Fig. 2). Tuning shifts were caused only by association between a tonal CS and an unconditioned stimulus because sensitization training was ineffective. Also, this plasticity was not caused by arousal to the CS frequency because RF shifts are still observed after training when subjects are placed under general anesthesia, a state lacking arousal to stimuli.
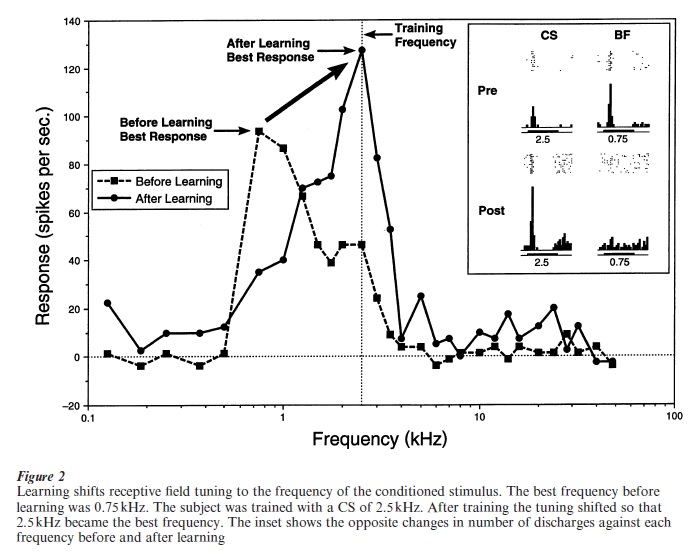
Several follow-up experiments showed that this neural plasticity was highly specific to the frequency of the CS; responses to tones a small fraction of an octave away were unchanged or decreased. Also, discrimination training (one tone reinforced, another tone not reinforced), produced increased responses to the former and decreased responses to the latter. Further, RF plasticity developed within only five trials of conditioning and could be retained as long as eight weeks after training. Moreover, instrumental avoidance training also produced the same shifts in tuning, both for one-tone training and for two-tone discrimination training. Habituation produced the opposite type of RF plasticity, a decrease in response that was highly specific to the frequency of the repeated tone. Finally, RF plasticity was found to exhibit a key characteristic of memory, that of consolidation, i.e., for periods of hours to days after a single training session, the RF plasticity became stronger and more specific. Thus, neural plasticity of receptive fields in the primary auditory cortex shares all of the major characteristics of associative memory; in addition to being associative, RF plasticity is highly specific, discriminative, develops very rapidly, is very long lasting and shows consolidation. Therefore, RF plasticity may be considered to be ‘physiological memory’ (Weinberger 1998).
RF plasticity has been reported in many laboratories and several species and similar effects have been reported in studies of brain metabolism in animals. Moreover, brain-imaging studies also have found RF plasticity in the human auditory cortex during learning (Morris et al. 1998).
5. Future Directions
The mechanisms of RF plasticity are unknown but are currently under investigation (Edeline 1999). There is strong evidence that the induction of RF plasticity involves the release of a modulatory transmitter, acetylcholine, into the auditory cortex from the nucleus basalis of the basal forebrain. Responses of its cells to a CS increase during conditioning. Moreover, pairing a tone with stimulation of the nucleus basalis produces the same type of tuning shifts observed during normal behavioral learning and pharmacological blockade of cholinergic muscarinic receptors in the auditory cortex prevents such tuning shifts.
The functional role of receptive field plasticity in the auditory cortex is also under investigation. One hypothesis is that it represents a memory code for the psychological importance of stimuli; that is, the brain stores the acquired significance of sensory experiences by increasing the number of neurons that become tuned to the relevant stimulus. Such a memory code could explain why one is more likely to hear one’s own name in a noisy room than a random name. Also, it could explain why the loss of memory in aging and brain degeneration is less severe for the most important memories of a lifetime. Alleviation of deafness by electrical stimulation of the inner ear (‘cochlear implant’) involves new learning and thus probably RF plasticity. Future studies of neural plasticity in the auditory cortex are likely to be directed toward its cellular and molecular mechanisms, other sensory systems, and psychological and behavior functions.
Bibliography:
- Edeline J M 1999 Learning-induced physiological plasticity in the thalamo-cortical sensory systems: A critical evaluation of receptive field plasticity, map changes and their potential mechanisms. Progress in Neurobiology 57: 165–224
- Morris J S, Friston K J, Dolan R J 1998 Experience-dependent modulation of tonotopic neural responses in human auditory cortex. Proceedings of the Royal Society (Series B) 265: 649–57
- Weinberger N M 1998 Physiological memory in primary auditory cortex: Characteristics and mechanisms. Neurobiology of Learning and Memory 70: 226–51
- Young R M 1970 Mind, Brain and Adaptation in the Nineteenth Century. Cerebral Localization and its Biological Context from Gall to Ferrier. Clarendon Press, Oxford, UK
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality