
Sample Neural Basis Of Dreaming Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. iResearchNet offers academic assignment help for students all over the world: writing from scratch, editing, proofreading, problem solving, from essays to dissertations, from humanities to STEM. We offer full confidentiality, safe payment, originality, and money-back guarantee. Secure your academic success with our risk-free services.
From the animistic dawn of consciousness to the modern psychoanalytic era, dreams have been central to the mythologies of human cultures. Because of their peculiar formal differences from waking consciousness, dreams have typically been thought to represent cryptic messages emanating from sources outside of the dreamer (such as the godhead and other disembodied spirits) or from within the self (such as unacceptable desires arising from the depths of one’s own unconscious mind). Dreams have thus been considered to be proverbial, privileged, and prophetic signals so deeply encoded as to require interpretation by a specialist. The interpretation of dreams has fallen to shamans, to priests, and since 1900, to psychoanalysts. But despite vigorous self-promoting appeals, not even the latter group could make any legitimate claim to a genuine science of dreaming. It is only within the second half of the twentieth century that sleep research has provided an objective, experimental foundation for the study of dreaming. By establishing a solid tie with neurobiology, it has also created the opportunity to understand this fascinating mental state in terms of brain physiology.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
In such a young field, the data and the conceptualizations of them are still naturally fragmentary, preliminary, and rapidly evolving. And it should be recognized that while animistic and external agency models can no longer be countenanced, a new psychology of dreaming that emphasizes undisguised personal meaning might find a scientifically legitimate place. In fact, it would appear that one of the important functions of dreaming could well be the reconfiguration of emotionally salient biographical information. If this were true then the interpretation of dreams might be quite simple: the dream is its own interpreter; its meanings are transparent; and its formal peculiarities are the straightforward expression of the unique information-processing mode of the brain in sleep.
1. Definition Of Dreaming
Dreaming is a distinctive mental state that occurs periodically in normal human sleep. Typical dream reports include such psychological features as hallucinations, delusions, cognitive abnormalities, emotional intensifications, and amnesia. These five remarkable features of dreaming have invited its comparison to abnormal states of mind that occur during waking in certain clinical conditions, especially delirium and the organic mental syndromes. The exploration of the neurobiological basis of dreaming therefore constitutes not only an aspect of mind-body interaction but also provides a model for understanding the brain basis of mental illness.
An obvious obstacle to the scientific study of dreaming is that the unconfirmable nature of all subjective experience is compounded by difficult access to the mind in sleep. The recent development of sleep laboratory recording techniques has given the study of dreaming an instrumental and systematic character, and the emerging data encourage psychophysiological integration. The constellation of psychological features of dreaming, their concomitance with objectively identifiable physiological states, and the occurrence of the same physiological states in all mammals allows the investigation of the neurobiology of dreaming in animal preparations.
1.1 Psychological Features Of Dreaming
Dreams are characterized by vivid and fully formed hallucinatory imagery with the visual sensory domain predominant; auditory, tactile, and movement sensations are also prominent in most dream reports. Compared with the common representation of these sensorimotor domains, taste and smell are underrepresented. Reports of pain are exceedingly rare despite the involvement of dreamers in frightening and even physically mutilating scenarios. Table 1 summarizes the formal features of REM (rapid eye movement) sleep dreaming.

Dreaming is properly considered delusional because subjects have virtually no insight regarding the true nature of the state in which they have their unusual sensorimotor experiences. Thus the tendency is great to consider dream events as if they were completely real during the dream even though they are promptly recognized as fabrications when recalled in subsequent waking states. This lack of insight is all the more surprising since uncritical belief in the reality of dream events must overcome high degrees of improbability and even physical impossibility.
The lack of insight that makes dreams delusional is part of a broader set of cognitive disturbances. Dreams are characterized by discontinuities (with unexplained changes of subject, action, and setting); by incongruities (with social inappropriateness and cognitive illogicality); by improbabilities and impossibilities (with defiance of physical law); and by uncertainties (with explicit vagueness). Dream characters and dream objects may be banal or they may be altogether fantastic and impossible collages of existing reality; they may behave normally or indulge in the most absurd, improbable, or impossible actions in settings either familiar or bearing only the faintest resemblance’s to those of real life. To explain these unique and remarkable dream features, illogical thought processes such as non sequiturs, post hoc explanations, and mythical, metaphorical, and symbolic elaborations are the norm.
Memory undergoes a paradoxical intensification and suppression during dreaming; remote recall is intensified within the dream as characters, scenes, events, and concerns from previous experience are knitted into the fanciful and evanescent fabric of the plot. Dreams can thus be said to be hypermnesic within the state itself. This increased access to memory within the state of dreaming contrasts markedly with the virtual impossibility of recovering the dream product after the state has terminated. On awakening, even from a dream in progress, subjects have difficulty holding the vivid experiences in short-term memory long enough to give a report or transcribe the dream scenario in detail. Thus there is amnesia for the hypermnesic dream. It can be conservatively stated that at least 95 percent of all dream mentation is entirely unremembered.
Emotion fluctuates widely in association with the abnormal and vivid mental content of dreaming; anxiety, elation, and anger are the three affects that undergo most marked intensification during dreaming. Obsessional concerns are common with dreamers focusing their worry on nudity, being lost, missing trains, and a host of other incomplete arrangements. Depressive affects are markedly under-represented with shame and guilt playing a relatively small part in dream emotion. Contrary to the predictions of psychoanalytic dream theory, sexual dreams are disappointingly rare.
Mental experience with some or all of these dream features can occur in any physiological sleep stage but it is most common at sleep onset, in REM sleep, and in late night light NREM sleep. Early in the night, deep NREM sleep is usually accompanied by little or no recallable mentation or by perseverative thinking about the individual’s ongoing events and concerns.
2. The Association Of Dreaming With REM Sleep
In 1953, Eugene Aserinsky and Nathaniel Kleitman noted that the sleep of children began and was periodically interrupted by activation of the electroencephalogram and by bursts of saccadic eye movement, the so-called rapid eye movements, or REMs. William Dement confirmed the hypothesis that these periods of brain activation during sleep were correlated with dreaming. The capacity to recall dreams appeared to be related to the nature of the awakening process: subjects who learned to obtain a fully aroused state without moving increased their recall capacity. Home-based recording methods, such as the Nightcap (see Ajilore et al. 1995) have recently confirmed these laboratory observations. Because they are simpler, less disruptive, and cheaper, they can greatly extend this data and put objective dream research within reach of any scientifically inclined psychologist, anthropologist, or social scientist.
Within each REM period, dream intensity tends to parallel the intensity of phasic physiological events, especially the eye movements; arousal during REM sleep with flurries of eye movement activity yielded reports fulfilling the definition of dreaming given here in 90–95 percent of the cases (some aspects of this physiology are shown in Fig. 1). When scored for vividness, emotionality, and imagined physical activity, these measures were correlated positively with the quantitative intensity of the eye movement in the REM sleep just prior to awakening.
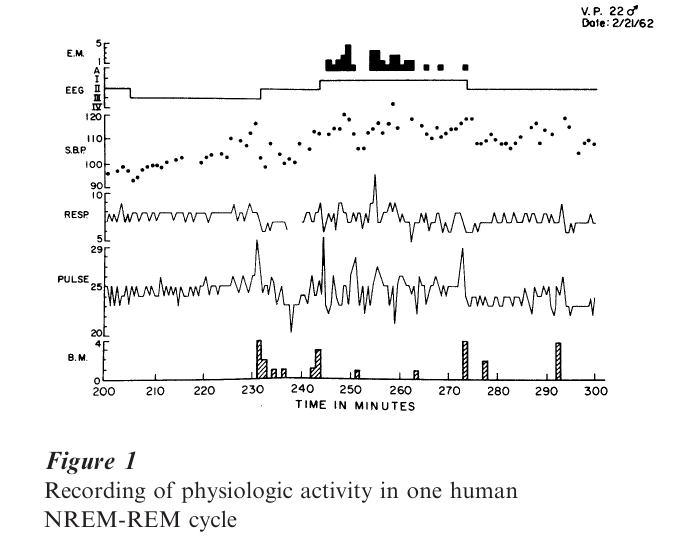
Estimations of dream duration correlate positively with the time spent in REM sleep prior to arousal. When subjects were aroused after only 5 minutes of REM sleep, they gave shorter reports; after 15 minutes had elapsed, reports were considerably longer and more detailed. Thus it would appear that despite expansion and contraction of temporal duration estimates within individual dream scenarios, the overall correlation between time estimation of dream duration and real time elapsed in REM sleep is quite good. This finding helps to dispel the skeptical notion that dreams occur in the instant before awakening or as part of the awakening process. To test the resistance of memory to dreams, awakenings were performed in the non-REM sleep phase at intervals following the termination of REM. The incidence of reported dreams dropped to non-REM levels within 5 minutes, indicating the extremely fragile state of memory within sleep and highlighting the strong state dependency of recall upon arousal from REM sleep.
3. A Psychophysiological Theory Of Dreaming
The simplest and most direct approach to the correlation of dream mentation with the physiological state of the brain is to assume a formal isomorphism between the subjective and objective data. By isomorphism is meant a similarity of form in the psychological and physiological domains. For example, the subjective experience of visually formed imagery during dreaming implicates activation of perceptual elements in the visual system in a manner formally similar to that of the waking state. Other details of psychophysiological correlation are assumed to obey the same general law; for example, the vivid hallucinated sensation of moving is assumed to be related to patterned activation of motor command systems and those central brain structures subserving the perception of position, and changes in position, of the body in space.
To be fully adequate, a psychophysiological hypothesis has to account for the following processes, which are illustrated in Fig. 2 and described in more detail in the entry on Sleep States and Somatomotor Activity.
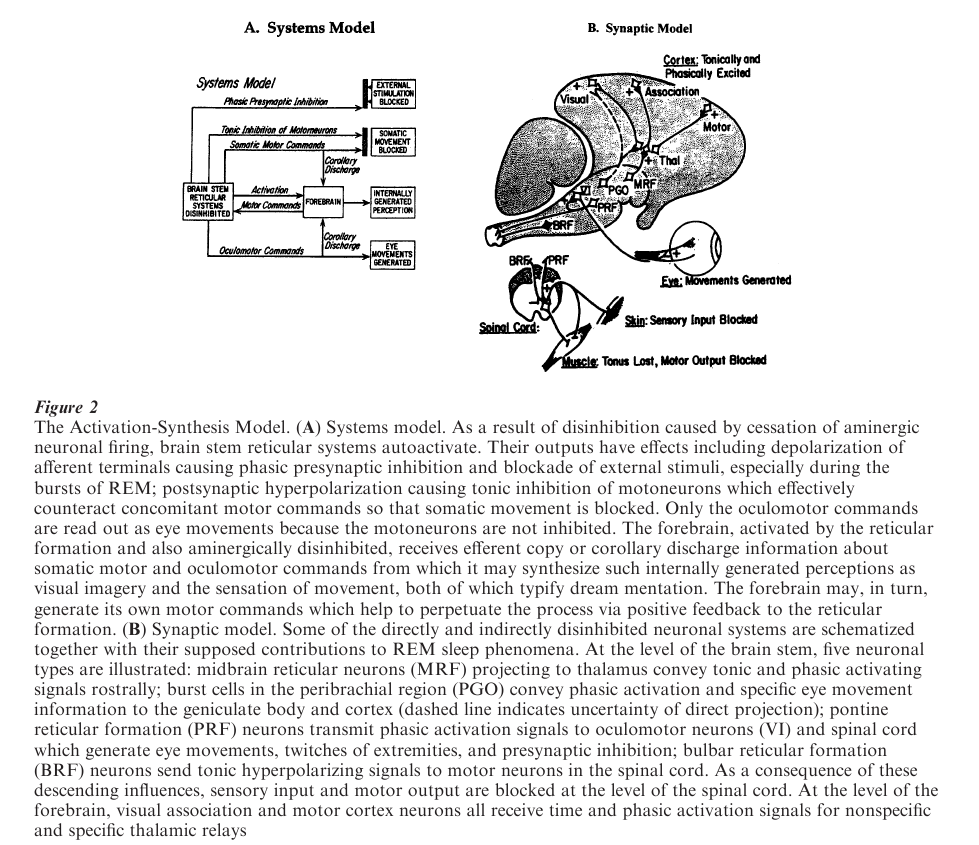
3.1 Activation
The brain has to be turned on and kept internally activated to support dream mentation throughout the REM sleep episode. A possible mechanism for this activation process is disinhibition of sensorimotor circuits (related to cessation of activity in aminergic inhibitory neurons) and internal excitation (owing to the reciprocal increase in cholinergic activation) Aminergic neurons decrease their output by 50 percent in non-REM sleep and virtually cease firing during REM while cholinergic neurons resume their high waking levels of discharge and fire in clusters as the eyes move back and forth.
3.2 Input Blockade
Input from the outside world to the internally activated brain has to be prevented in order for sleep and the illusions of dreaming to be maintained. This is accomplished in at least two ways. One is presynaptic inhibition of the afferent terminals of the group-Ia afferent nerves, which impedes access of external stimuli to the central nervous system. Presynaptic inhibition has also been recorded at secondary relay nuclei throughout the brain stem and thalamus guaranteeing even more complete insulation of the activated brain from the outside world. The second mechanism for excluding sensory input is the occlusion by internally generated messages that occupy the higher levels of brain sensory and perceptual circuits. Rodolfo Llinas and Denis Pare theorize that the deafferentation of REM sleep is a major determinant of the difference between dreaming and waking consciousness.
3.3 Output Blockade
The internally activated and aminergically deafferented brain must also quell motor outputs to prevent the disruption of sleep by reafferent stimulation and to halt the enactment of dreamed motor commands. This is accomplished by postynaptic inhibition of final common path motor neurons in the spinal cord. The dreamer who fancies himor herself walking, running, swimming, or flying is, in fact, paralyzed from the neck down.
3.4 Internal Signal Generation
It remains to provide the activated but sensorially disconnected brain with internal signals, which it can then process as if they came from the outside world. This appears to occur in part by a mechanism intrinsic to brain activation: the reciprocal interaction of aminergic and cholinergic neurons in the brain stem. In most mammals including humans, the so-called PGO waves (P, pons; G, (lateral) geniculate; and O, occipital cortex), present themselves as candidates for an internally generated information signal arising at the level of the pontine brain stem. In association with oculomotor activity, strong pulses of excitation are conducted by reticular pathways to the thalamus and radiated by independent pathways to the visual and association cortices.
It is now known that the PGO waves are generated by cellular activity, which faithfully replicates the directional aspects of generated eye movements at the level of the brain stem. Thus not only is internal information generated but the information has a high degree of spatial specificity. According to the activation-synthesis hypothesis of dreaming, the now autoactivated and autostimulated brain processes these signals and interprets them in terms of information stored in memory. Recent evidence indicates that the PGO system is facilitated by cholinergic and restrained by aminergic neuromodulation.
Three important observations concern the origin and fate of motor command signals arising in the brain during REM sleep.
The first observation, suggesting that the motor commands would generate behavior if not inhibited, is the revelation by Michel Jouvet and by Adrian Morrison of elaborate motor behaviors during REM sleep in cats that have lost their capacity to inhibit muscle tone; these behaviors include running, pouncing, attack and defense postures, and intense fear reactions accompanied by hissing and piloerection.
The second observation, suggesting a brain stem origin of the motor patterns generated in REM, is that the locus for the elicitation of locomotion by electrical and chemical stimuli is immediately adjacent to the cellular controllers of REM in the anterodorsal pontine brain stem.
The third observation, offering dramatic proof that brainstem motor commands are actually the building blocks of dream plots, is the emergence of dream content specific movements in human subjects who have lost their ability to inhibit motor activity in sleep owing to degenerative disease of the brainstem.
4. The Neuropsychology Of Cognitive Deficits In Dreaming
Although, the basis of the cognitive disturbances occurring in dreaming is not yet well understood, it is possible that the deficits are related to the cessation of activity in aminergic neurons. The REM sleep arrest of aminergic neuronal activity affects the entire brain including the cerebral cortex by depriving it of the tonic modulatory influence of both norepinephrine and serotonin. This tonic aminergic modulatory influence may be essential to waking attentional processes, including the capacity to organize information in a logical and coherent manner and to achieve full self-awareness. In the absence of external cues and deprived of internal modulatory influences by aminergic systems, the activated forebrain thus could interpret its internally generated signals as if they were external stimuli.
By a similar mechanism it may be speculated that the synthesized dream product is unremembered. That is, the activated forebrain circuits which mediate the dream experience are simply not instructed to keep a record of the transaction. The mnemonic instruction is also postulated to be carried out by signals from the aminergic interneurons to their vast postsynaptic domain in waking (when they are active) but not in dreaming (where they are inactive). Current models of learning and memory at the cellular level evoke the intervention of an aminergic interneuron. The attribution of dream amnesia to the loss of aminergic modulation is consistent with these hypotheses.
The complementary results of recent positron emission tomography (PET) imaging and brain lesion studies of dreaming have greatly enriched the experimental study of dreaming in humans. These results are summarized in Table 2.

Two forebrain regions appear to be critical for the maintenance of dreaming. One is the parietotemporal-occipital-cortex, a region known to be important to visuomotor imagery. The other is a subcortical zone connecting the limbic emotion centers with the frontal cortex. PET studies indicate that both of these regions are selectively activated in REM sleep, and when either is damaged by a brain lesion, subjects report a marked dimunition in the subjective experience of dreaming. The PET studies also indicate that while emotion centers in the amygdala and related limbic cortical regions are turned on, the dorsolateral prefrontal cortex is selectively deactivated in REM. This reciprocal pattern maps on to the intensification of emotion and to the deficiencies in cognition that typify dreaming.
5. Summary
The activation-synthesis model of dreaming and the reciprocal interaction theory of sleep cycle control on which it is based are both incomplete and controversial models. They represent linked working hypotheses about the fundamental physiology of sleep and the way in which that physiology may help us to understand unique features of the dream process. The attribution of automaticity to the control system and the feature of randomness in the information generator model should not be taken to exclude the meaningful nature of the synthetic process carried out by the activated dreaming brain. The brain–mind of each dreamer is obliged to make as much sense as it possible of its internally generated signals under the adverse working conditions of REM sleep. Thus, the dream product of each individual may reveal unique stylistic psychological features and emotional concerns that are worthy of scrutiny by individuals interested in reviewing life strategies. But the new theory challenges the psychoanalytic idea that the many meaningless aspects of dream mentation are the result of an active effort to disguise the meaning of unconscious wishes, which are in turn postulated to be the driving force of dreaming. Instead, it ascribes these defective cognitive properties of dreaming to unusual operating features of the internally activated, auto-stimulated brain during REM sleep.
Bibliography:
- Ajilore O A, Stickgold R, Rittenhouse C, Hobson J A 1995 Nightcap: Laboratory and home-based evaluation of a portable sleep monitor. Psychophysiology 32: 92–8
- Antrobus J 1991 Dreaming: Cognitive processes during cortical activation and higher afferent thresholds. Psychological Reviews 98: 96–121
- Hobson J A 1988 The Dreaming Brain. Basic Books, New York
- Hobson J A, Pace-Schott E, Stickgold R 2000 Dreaming and the brain: toward a cognition: Neurosciences of conscious states. Behavioral and Brain Sciences 23: 793–842
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality