
Sample Motor Control Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
The rich varieties of human and non-human animal movement are the final product of motor mechanisms within the nervous system and the muscles they innervate. The selective contraction and relaxation of these muscle groups support the organism (posture), mediate internal functions such as heartbeat, breathing, and peristalsis of the digestive system, and propel the creature through its external world. The motor neurons typically work in concert with interneurons and sensory mechanisms that are orchestrated together through intricate patterns of activity. The motor control pathways in whole organism actions read-out these patterns from many parts of the brain and its contact with the sensory world. It has been argued (Lashley, 1951 Sperry 1952) that movement, and thus motor control, is the most fundamental property of nervous systems. Without movement there is no behavior as the term is normally defined. Movement also provides overt and objective measures of the rich symphonies of behavior we see in the natural world.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Commonalities And Specializations In Mechanism
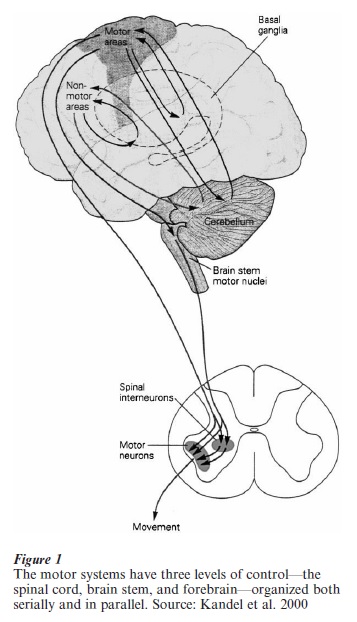
Motor control in all vertebrate species (animals with backbones) operates from a similar ground plan, but with important differences in detail. For all vertebrates spinal and brain stem mechanisms mediate a number of basic forms of movement, such as locomotion and postural adjustment. In mammals, and especially in humans, higher cortical areas along with the basal ganglia and cerebellum contribute importantly to the acquisition, planning, flexibility, and fine-tuning of movement sequences (Fig. 1). Major properties of these structures are discussed below. In invertebrates (animals without backbones) the basic building blocks of motor control are diverse and distinct from those found in vertebrate species, yet there is a remarkable convergence in terms of overall solutions to motor control. For example, relatively small groups of neurons, and in some species single cells, generate rhythms of an endogenous (intrinsic to the cells) rhythms without the need for molding by afferent influences. These central pattern generators were first documented clearly in invertebrates, and then determined for vertebrates (Fentress 1976, Pearson 1993; see Comparative Neuroscience). In the intact animal these patterns of spontaneous activity are typically modulated by other neural and endocrine influences, however.
Most investigators today believe that many of the most important questions of motor control rest at the interface between intrinsic properties of neurons and their sensitivity to extrinsic factors. Motor commands may remain latent until triggered by descending influences, rather as dropping a needle onto a phonograph record triggers patterns of sound programmed within the chosen record. The analogy to music turns out to be a useful one, for it reminds us that we wish to account for the notes (basic units) of movement, the construction of chords of simultaneously activated motor elements, melodies in time, rhythmic principles, volume (intensity) and timbre (fine tuning), and even possible transpositions from one moment to the next. Questions of movements guided toward specific features of the environment are also of obvious importance, and are discussed in other chapters.
2. Early Models And Current Perspectives
Early models of vertebrate movement stressed individual reflexes and reflex chains as the basic building blocks in the organization of action patterns (Sherrington 1906). Spinal reflexes operate through relatively simple chains of neurons, where sensory and interneurons link up with motor neurons that in turn project to a select group of muscles. Interest in reflex behavior (such as the well-known knee jerk reflex elicited by your local physician) remains intense, and reflexes are clearly essential for the production of posture, basic movements, and defensive reactions. Their speed and reliability of production reflect what are often called ‘automated’ properties of motor control, that is movement profiles that are rapid, essentially ballistic, and not mediated by conscious processes. Even these reflexes can be modulated by higher neural activities, however. As in nearly all aspects of neural function it is thus a matter of looking at intrinsic circuits together with extrinsic influences. These occur at many levels of neural control.
2.1 Movement Units And Their Limits
One of the basic challenges in the study of motor control is the dissection of fundamental movement units. In this terminology a unit is a relatively invariant pattern of muscular contractions that are typically elicited together. Reference to one of the most complex forms of human movement control, speech, is illustrative (Neville 1995). When we speak we articulate a broadly invariant set of phonemes (basic sound patterns) in sequence. This allows for the articulation of speech to be relatively ‘thought free’ in normal discourse, and allows the listener to decode the message through a complex array of perceptual and higher-order cognitive processes. The example is useful in that in the smooth flow of speech individual phonemes are often ‘co-articulated’ with the fine details of one sound unit reflecting the production of preceding and subsequent sound patterns. So even here the ‘intrinsic’ properties of a speech unit can be sensitive to extrinsic influences as defined by other speech units. This is what gives speech its smooth and flowing nature (as opposed, for example, to most computer generated ‘speech’).
Thus, in some sense movement and their motor control ‘units’ are abstractions that frequently blur at the edges, and which can be defined from a number of complementary perspectives (Golani 1992). In complex forms of movement, such as playing a sport or musical instrument, or dancing with a moving partner, many neural pathways are orchestrated together in dazzling patterns that are widely distributed, serially ordered, and parallel in their operations. It is for this reason that the very distinction between motor control systems and other properties of the nervous system are often difficult to untangle.
2.2 Movement Notation
A number of movement notation systems have helped clarify the organization of motor patterns at many different levels. For example, the Eshkol–Wachman movement notation system (Golani 1992) has helped investigators separate specific parameters of movement, such as movements in a plane (flat surface) and those that involve rotations or conical movements around a single limb. These notational systems, derived from human choreography, also remind us that movement is in its essence both relational and dynamic. Its description also depends upon the frame of reference adopted. For example, we can move our bodies with respect to internal coordinates and yet maintain a fixed relationship to external events, such as a moving social partner. Often the units of control involve multiple muscular systems, such as when we produce relatively invariant motions during the final phases of feeding or brushing our teeth via complementary adjustments of individual muscular acts (such as those involved in arm movements or turning the head).
2.3 Dynamic Systems
Many workers today have chosen to supplement the language of motor units and experimentally isolated mechanisms with insights derived from dynamic systems theory (Kelso 1995, Thelen 1985). These approaches remind us that behavior (movement, and thus motor control) is fundamentally imbedded in time, and involves the cooperation of a rich multitude of processes whose boundaries are often difficult to define. Further, there has been an increased appreciation for the flexibility of underlying mechanism operations. Even in invertebrates circuits that contribute to a given movement may be shared and change their internal properties from one occasion of expression to the next (Marder and Calabrese 1996). This ‘remolding’ of circuit properties adds importantly to the challenges of deep understanding of the processes of motor control.
3. Diversity And Plasticity In Human Movement
The human species is especially remarkable in its ability to master skills of an enormous diversity. One needs to reflect just for a moment on actions such as speech, sports, dance, driving an automobile, or musical performance to appreciate this fact. Indeed while each species is the master of its own behavioral niche, it is in humans that we find the most rich variety of forms of expression. Our nervous systems, throughout much of our lives, are remarkably plastic, meaning that they can be refined and remolded into an often dizzying array of specialized talents.
We have all had the experience that in the early learning of a new motor skill our attention is fully absorbed. Later we produce the actions in an automated manner, without any conscious awareness. When we seek to modify this skill, or apply it to a new context, the Executive parts of our nervous system are recaptured, with thought and planning once again playing a major role. It is now well established that different neural circuits are employed in the early and late stages of skill acquisition (Posner et al. 2001).
3.1 Basic Pathways
It remains possible to separate particular neural pathways that contribute to different properties of movement. The human motor system can be visualized in part as being composed of descending pathways derived from the cerebral cortex and brainstem that are fundamental to the suprasegmental control of movement. The term ‘segmental’ refers to spinal segments that control different muscle groups in an orderly way. Suprasegmental thus refers to actions that incorporate muscles of different extremities such as the head, arms, trunk, and legs. The basal ganglia and cerebellum contribute to the planning, sequencing and fine-tuning of movement. The basal ganglia are embedded deep within the forebrain and also include midbrain pathways. These pathways do not connect directly to motor pathways, but exert important indirect influences that are mediated through cortical and thalamic loops. Disorders of the basal ganglia greatly compromise effective movement, such as seen in Parkinson’s Disease with its characteristic tremor and poverty of movement, and Huntington’s Disease with its characteristic choreoform (sporadic and ballistic) movements. Each of these diseases is also marked by cognitive (thought, planning) deficits as the disease progresses, highlighting once again how motor systems are intimately linked with other neural functions.
The cerebellum has multiple sensory and motor functions, among which the timing and fine-tuning of motor control are among the most apparent. It is now known that lesions of the lateral cerebellum also compromise the perception of temporal events (Ivry 1996), thus illustrating yet again how distinctions between motor and other neural functions can be misleading. Individual circuits, and even individual neurons, can mediate a variety of functions.
The spinal cord, which receives descending inputs from the brain, is neatly organized with anatomically arranged segments influencing and controlling actions of the upper and lower body. These richly interconnected pathways work together with the neuromuscular system which is also composed of nuclei of the motor cranial nerves and the muscles that they innervate.
At every level of organization major afferent pathways project to the motor control mechanisms, but their influence is a function of such factors as speed of movement and the extent to which movement patterns have become automated (as in well-practiced tasks). Often the influence of afferent pathways is relatively small in well-practiced and simple automated movements, but sensitivity to sensory influences can attain major importance when actions have to be adjusted, such as in the confrontation with unexpected circumstances.
Activities from extrinsic sources are gated to varying degrees as a function of the demands placed upon the organism at the moment. Routine activities run more or less ‘autonomously,’ but are able to be corrected if circumstances change. In this way the production of movement sequences can be both automated and responsive to novel circumstances. There are many known human motor diseases that can selectively affect both the automated performance of movement and the ability of the individual to adjust to novel situations (Linas 2001). A skillful neurologist can, by examining detailed properties of movement performance, often pinpoint parts of the nervous system that are damaged.
Much of what we call voluntary movement (as opposed, to the elicitation of simple reflexes) is mediated by connections that in a sense ‘wander’ through a complex neuronal maze from the cerebral cortex to the spinal cord. Thus, the primary motor cortex, which is somatotopically organized as is the somatic sensory cortex (meaning that different body parts are connected to specific neural regions), influences specific muscle groups. The cortex itself is layered, with neurons forming groups that are rather like a series of pies stacked upon one another. It is in what anatomists call layer V (counting from the top of the cortex) of the primary motor cortex that one finds projections directly to motor neurons and interneurons, in the ventral (‘bottom’) level of the spinal cord, via what is called the corticospinal tract. In humans there are approximately one million axons (those processes of nerves that carry signals out from the neuron’s cell body) and just under 50 percent of these originate from the motor cortex. Other parts of the cortex, such as the premotor areas and the supplementary motor areas, also exert important and distinct influences upon our movement skills. Thus our cortices, the highest parts of the human brain, connect us more or less directly to the world through movement.
From the cortex the neural axons pass through white matter of the sub-cortex, through the internal capsule and cerebral peduncles on their way to the spinal cord, and hence the muscles that make movement possible (see Fig. 1). The term ‘white matter’ refers to the fact that these neurons have a fatty insulating sheath that promotes rapid and reliable transmission of signals from one neuron to another. These tracts in turn form the medullary pyramids deep within the brain (a region called the medulla). The projection encompasses what has classically been called the pyramidal tract. In addition to the pyramidal tract there is the extrapyramidal system, which is composed of other nuclei, such as those found in the basal ganglia. These extrapyramidal systems are for the most part more indirect in their motor control functions than are the pyramidal pathways, but their importance should not be underestimated. For example, linkages between emotional and cognitive processes and the motor system are largely mediated through these extrapyramidal systems.
The left side of the brain primarily is involved with movements on the right side of the body, and vice versa, but the crossing of fibers is incomplete. Ultimately, the coordination of movements from each side of the body involves unified actions of neural circuits that are in themselves primarily concerned with one body side.
The corticospinal tract itself makes monosynaptic (i.e., direct) connections with motor neurons, which in turn control muscular contractions. There are also many connections to interneurons in the spinal cord. The importance of these indirect connections is that they allow a number of influences from other brain regions that are essential for adaptive motor performance. Throughout, the activities of the corticospinal tract and related pathways incorporate a rich variety of descending modulating influences.
3.2 Movement Disorders And Development
In addition to the often tragic consequences of neural damage (e.g., stroke) the neurologist and behavioral neurobiologist can gain important insights into the foundations of motor control by examining the developmental progression of motor skills. There are predictable milestones in the construction of human motor skills. A relatively recent perspective has stressed the theme of ‘ontogenetic adaptations’ in motor control (Oppenheim 1982). In the past it was customary to think of nervous systems starting out as ‘imperfect’ adult systems—rather like a linear progression from less than fully adequate to fully adequate. This view has now changed. Think, for example, of the newborn infant who must swallow (which involves a complex coordination of 20 plus muscles in the throat; Doty 1976), and breathe and suckle. There is no room for error or practice. Indeed, many motor circuits are formed in a prefunctional manner. This means that the circuits are established without the benefit of overt motor practice. At a more detailed neuronal level these circuits develop in what has been called a retrograde manner, with the establishment of final neuromuscular and other motor circuits preceding the establishment of higher-order interneuronal influences and thus responses to sensory events from the internal and external environments (Fentress and Gadbois 2001).
This reminds us of the work of ethologists (e.g., Hinde 1970, Lorenz 1950, Tinbergen 1963) who stressed the ‘instinctive’ nature of many actions in animals and humans. The basic idea is that many of the fundamental properties in animal and human behavior are a relatively direct reflection of our genetic make-up. People, for example, speak; other species do not. Many basic human and animal capacities for movement emerge relatively independently of specialized experiences. In some deep sense we are indeed ‘preprogrammed’ to be human, just as the bird, and bat, and insect are preprogrammed to be members of their own species. What is perhaps most remarkable about humans is that our genetic structure allows for enormous flexibility in the detailed organization of our movements. In that respect we stand alone among the animal species. Yet, for all animal species the delicate interplay between genetic and experiential factors (broadly defined, across many levels of organization) remains a major area of importance for further investigation. An essential point is that even the most basic processes of differentiation in development involve genes and experience working in concert (Hinde 1970, Edelman 1987).
How is it that we learn motor skills, and how are these skills mediated? At the present time our knowledge of these essential questions is very limited. Certainly many of the mechanisms we have touched upon contribute richly. Thus, cerebellar circuits are important in the establishment of newly acquired reflexes as well as more complex cognitive skills (Thompson, Thach). The basal ganglia as well as cortical mechanisms also contribute to perfections of motor performance through experience (Graybiel). The importance of cortical mechanisms is perhaps predominant, but a challenge for future research is to determine how various neural mechanisms operate together at various stages of skill acquisition and performance.
4. The Need To Combine Mechanistic And Systems Perspectives
A challenge of increasing importance in both the neural and behavioral sciences is how to capture the dynamic as well as modular properties of motor performance. Excellent overviews that cross-complementary levels of organization can be found in Gazzinga 2000, Kandel et al. 2000. Most investigators in the neurobehavioral sciences today take the position that any level of analysis must make contacts with levels that are both more molar and more molecular.
5. Challenges For Future Research
Human motor control systems are awesome in their richness. It is through these motor control systems that we interact as individuals, yet individuals deeply connected to other human individuals and to our animal brethren by solutions that evolution has enabled us with. We talk, sing, and dance. In spite of recent progress at many levels of research, precisely how we do these basic things remains deeply mysterious. Yet, piece by piece, we are beginning to understand the operation of individual mechanisms and their coordinated actions. For example, the rapid progress in brain imaging techniques has helped us understand how the final processes of motor control are connected to higher-order cognitive functions. The further understanding of motor control processes, including not just their performance on a short-term basis, but also their development and evolution, offers riches into the nature of the human psyche itself.
In recent years a number of model systems for motor control have been offered. One of the most promising is that there are variable foci of activity that occur in conjunction with variable expanses of collateral inhibition, that is, those events that block competing activity (Fentress 1976). For example, both the cerebellum and basal ganglia have lateral inhibitory (collateral) circuits that suppress action properties that would otherwise interfere with the primary focus of adaptive movement. There is now growing evidence that these collateral pathways can expand in their extent of influence as the core of motor performance becomes more tightly focused (Fentress 1976). What this means is that systems in the control of movement can be both broadly and much more finely tuned as circumstances dictate. Extrinsic influences are either accepted or rejected as the needs of the organism change.
In terms of the establishment of motor behavior during development, a number of genetic analysis that focus upon specific neural circuits are now being combined with experiments designed to clarify the roles and limits of specific sources of ontogenetic experience. The old ‘nature vs. nurture’ debate is now considered passe by most active investigators; the challenge is to see how genes and experience work as partners during the construction of brains and their behavioral manifestations.
In motor control we have much to learn about the detailed operation of specific mechanisms, but equally much to learn how these specific mechanisms become established and integrated into the rich symphonies of human and animal action. Whatever the details that will be revealed by future research, it is clear that motor control mechanisms not only reflect what we mean by behavior, but also give us insights into the inner workings of nervous systems that remain a major source of inspiration and mystery for all those who are fortunate enough to study them.
Major components of the human motor system. Spinal cord, brain stem, and motor areas of the cerebral cortex work together with direct and indirect connections of the basal ganglia and cerebellum in the planning and execution of integrated movement patterns.
Bibliography:
- Doty R W 1976 The concept of neural centers. In: Fentress J C (ed.) Simpler Networks and Behavior. Sinauer Associates, Sunderland, MA, pp. 251–65
- Edelman G M 1987 Neural Darwinism: The Theory of Group Selection. Basic Books, New York
- Fentress J C 1976 Dynamic boundaries of patterned behavior: Interaction and self-organization. In: Bateson P P G, Hinde R A (eds.) Growing Points in Ethology. Cambridge University Press, Cambridge, UK, pp. 135–69
- Fentress J C, Gadbois S 2001 The developments of action sequences. In: Blass E (ed.) Developmental Psychobiology. Kluwer Plenum, New York, pp. 403–41
- Gazzaniga M S (ed.) 2000 The New Cognitive Neurosciences, 2nd edn.
- Golani I 1992 A mobility gradient in the organization of vertebrate movement: The perception of movement through symbolic language. Behavioral and Brain Sciences 15: 249–66
- Graybiel A M 1995 Building action repertoires: Memory and learning functions of the basal ganglia. Current Opinion in Neurobiology 5: 733–41
- Hinde R A 1970 Animal Behavior, 2nd edn. McGraw-Hill, New York
- Ivry R B 1996 The representation of temporal information in perception and motor control. Current Opinion in Neurobiology 6: 851–7
- Kandel E R, Schwartz J H, Jessell T M 2000 Principles of Neural Science, 4th edn. McGraw-Hill, New York
- Kelso J A S 1995 Dynamic Patterns: The Self-organization of Brain and Behavior. MIT Press, Cambridge, MA
- Lashley K S 1951 The problem of serial order in behavior. In: Jeffries L A (ed.) Cerebral Mechanisms in Behavior. John Wiley and Son, New York, pp. 112–36
- Llinas R R 2001 I of the Vortex: From Neurons to Self. MIT Press, Cambridge, MA
- Lorenz K Z 1950 The comparative method in studying innate behavior patterns. Symposia of the Society for Experimental Biology 4: 221–68
- Marder E, Calabrese R L 1996 Principles of rhythmic motor pattern generation. Physiological Reviews 76: 687–717
- Neville H J 1995 Developmental specificity in neurocognitive development in humans. In: Gazzinga M (ed.) The Cognitive Neurosciences. MIT Press, Cambridge, MA, pp. 219–31
- Oppenheim R W 1981 Preformation and epigenesis in the origins of the nervous system and behavior: Issues, concepts and their history. Perspectives in Ethology 5: 1–100
- Pearson K G 1993 Common principles of motor control in vertebrates and invertebrates. Annual Review of Neuroscience 16: 265–97
- Posner M I, Rothbart M K, Farah M, Brucer J (eds.) 2001 The developing human brain. Developmental Science 4: 253–387
- Sperry R 1952 Neurology and the mind–brain problem. American Scientist 40: 291–312
- Thach W T 1996 On the specific role of the cerebellum in motor learning and cognition: Clues from PER activation and lesion studies in man. Behavioral and Brain Sciences 19: 411–31
- Thelen E 1985 Developmental origins of motor coordination: Leg movements in human infants. Developmental Psychobiology 18: 1–22
- Tinbergen N 1963 On aims and methods of ethology. Zeitschrift fur Tierpsychologie 20: 410–33
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality