
Sample Neural Basis Of Recognition Memory Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
The term recognition refers to the knowledge or feeling that an object has been encountered previously. Two different mental processes may contribute to recognition judgments: a general feeling of ‘familiarity’ and a specific ‘recollection.’ These two aspects of recognition may depend on different brain systems, but are treated together here.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Neural Systems Mediating Visual Recognition
For most sensory modalities (e.g., vision and touch), receptors convert sensory information into neural signals, and modality-specific cortical fields, such as the primary visual cortex (‘V1’ in Figure 1), process that information further. These early sensory processing stages are a prerequisite for recognition memory. However, the core recognition system must function more generally. If a blind person fails to recognize a visually presented object, we say that he or she has a visual impairment, not a failure of recognition memory. Therefore, this research paper will focus on general aspects of recognition and its neural basis, neglecting modality-specific ones.
1.1 Methods For Assessing Recognition Memory In Nonhuman Primates
Because both human and nonhuman primates rely heavily on visual information in their daily activities, recognition is best understood for visible objects. Visual recognition memory in monkeys has been studied by making selective brain lesions and by disconnecting one brain region from another. These studies identify whether a given brain structure is critical for recognition. Monkeys, of course, cannot tell us whether they remember seeing an object. For this reason, methods have been developed that allow them to reveal that information. One commonly used method is the delayed nonmatching-to-sample task (DNMS). In DNMS, each trial has two parts: a sample presentation followed by a choice test. During the sample presentation, the monkey sees a single object, which covers the central well of a three-well test tray. The monkey then displaces the object to obtain a small piece of food hidden in the well underneath. A few seconds later the monkey sees the same, nowfamiliar object plus a novel one, with one object over the left well of the test tray and the other over the right well. The monkey can obtain more food by displacing the novel object but not by pushing aside the familiar one. Thus, on the choice test, the monkey solves the problem by applying a ‘nonmatching’ rule, i.e., by choosing the object that does not match the sample. If the monkey consistently chooses the novel object on the choice test, one can infer that the monkey recognizes the sample. In the laboratory, there are many variations of this basic method. For example, monkeys can be trained using a ‘matching’ rule (the delayed matching-to-sample task, or DMS) rather than the nonmatching one. It is also common to use a computerized apparatus that presents two-dimensional pictures on a touch-sensitive video screen rather than using actual objects on a test tray.
1.2 Role Of The Medial Temporal Lobe In Monkeys
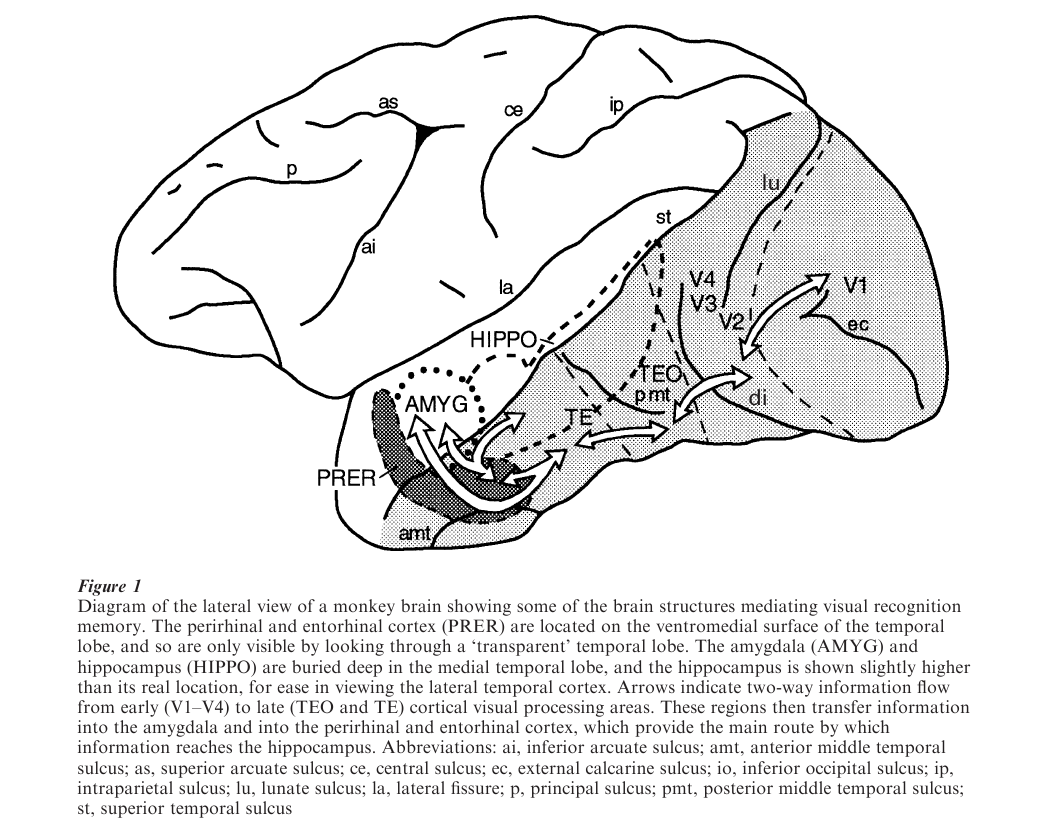
It has been known for many years that a limited set of brain structures, including components of the medial temporal lobe (MTL), are necessary for storing information about facts, objects, and events from all sensory modalities (Squire 1987). Early investigations into the neural basis of recognition memory pointed to two structures buried deep in the MTL: the amygdala and hippocampus (Mishkin 1982, see Figure 1). However, those experiments involved the mechanical removal of brain tissue, a technique that produced unintended damage to other parts of the MTL. More recent studies have used stereotaxic injection of neurotoxins to make more selective tests. These experiments show that, within the MTL, the perirhinal cortex and the entorhinal cortex, rather than the amygdala and hippocampus, are essential for visual recognition (Murray 2000). Monkeys with an intact amygdala and hippocampus, but without the perirhinal and entorhinal cortex, cannot recognize objects if they have to remember them for more than a few seconds.
Experts accept a central role for perirhinal and entorhinal cortex in object recognition and also agree that the amygdala is not important for this type of memory. Controversy remains, however, about the potential contribution of the hippocampus. Damage to the hippocampus leads to impairments in recognition memory in some studies but not in others. For example, one test of recognition relies on the tendency of monkeys to spend more time looking at novel objects than at familiar ones. This innate tendency is called ‘preferential viewing.’ Although hippocampal damage fails to produce consistent and systematic deficits on DNMS, it does disrupt preferential viewing. This could be because the preferential viewing test, unlike DNMS, does not rely on learned rules, or because the stimuli have no association with food. Interestingly, on DNMS, larger perirhinal and entorhinal cortex lesions cause greater memory deficits than smaller ones, which is what one would predict if a structure were playing a direct role in a particular function. The reverse relationship holds for the hippocampus. That is, larger hippocampal lesions are associated with better recognition memory than smaller lesions, suggesting an indirect relationship of the damage to recognition memory deficits. The fact that only the perirhinal and entorhinal cortex have the predicted relationship to recognition memory supports the view that they are the parts of the MTL mediating that function.
Figure 1 Diagram of the lateral view of a monkey brain showing some of the brain structures mediating visual recognition memory. The perirhinal and entorhinal cortex (PRER) are located on the ventromedial surface of the temporal lobe, and so are only visible by looking through a ‘transparent’ temporal lobe. The amygdala (AMYG) and hippocampus (HIPPO) are buried deep in the medial temporal lobe, and the hippocampus is shown slightly higher than its real location, for ease in viewing the lateral temporal cortex. Arrows indicate two-way information flow from early (V1–V4) to late (TEO and TE) cortical visual processing areas. These regions then transfer information into the amygdala and into the perirhinal and entorhinal cortex, which provide the main route by which information reaches the hippocampus. Abbreviations: ai, inferior arcuate sulcus; amt, anterior middle temporal sulcus; as, superior arcuate sulcus; ce, central sulcus; ec, external calcarine sulcus; io, inferior occipital sulcus; ip, intraparietal sulcus; lu, lunate sulcus; la, lateral fissure; p, principal sulcus; pmt, posterior middle temporal sulcus; st, superior temporal sulcus
The same parts of the MTL that are essential for visual recognition memory subserve tactual recognition memory as well. They are also critical for many types of stimulus–stimulus association memory. Accordingly, their function may best be characterized as one of object identification. Through this process, the visual, tactual, gustatory, olfactory, and auditory properties of objects are somehow linked or ‘bound.’ Thus, the perirhinal and entorhinal cortex are essential not only for visual recognition, but also for storage of information about objects generally (Murray 2000).
1.3 Role Of The Thalamus And Frontal Lobe In Monkeys
The perirhinal and entorhinal cortex do not operate alone, but instead work in concert with other anatomically connected brain structures that, together, form a neural network mediating visual recognition memory. At least two other brain regions participate: the medial thalamus and the ventromedial part of the prefrontal cortex (Mishkin and Murray 1994). Damage to either of these structures produces essentially the same effect on visual recognition memory as does damage to the perirhinal and entorhinal cortex. In addition, the thalamic and frontal regions noted above are connected with each other, and each is connected to the perirhinal and entorhinal cortex. These structures need to interact to mediate visual recognition; surgical manipulations that prevent information from one structure from reaching a second produce a deficit in recognition memory much like that seen after lesions to each individual site (Parker and Gaffan 1998). Precisely how these structures interact to produce recognition memory remains uncertain; it is possible that each structure underlies some unique aspect of recognition memory.
1.4 The Recognition Network In The Human Brain
The same neural structures that are critical for recognition memory in monkeys appear to be essential in humans as well. For example, memory impairments are found in humans following damage to the MTL as a result of either brain infections (e.g., Herpes encephalitis) or temporary interruption of the blood supply. Damage to the medial thalamus, as a result of Korsakoff’s disease, and to portions of the ventromedial prefrontal cortex, as a result of aneurysms of the anterior communicating artery, yield similar effects. It must be kept in mind, however, that in clinical cases brain damage rarely respects the boundaries of individual structures, so that the kind of precision outlined above for monkeys is not often possible for the human brain. Nevertheless, there is evidence that in humans, as in monkeys, the perirhinal and entorhinal cortex are important for recognizing objects. And, as in monkeys, there is controversy about the role of the hippocampus. People with damage to the hippocampus and adjacent perirhinal and entorhinal cortex are much more impaired in stimulus recognition than those with damage restricted to the hippocampus. Whereas some studies have shown that humans with damage limited to the hippocampus and closely related structures have relatively intact recognition abilities (Aggleton and Brown 1999), others have reported that such subjects are, on average, impaired in recognition memory (Squire and Knowlton 2000). In humans, the hippocampal contribution to recognition memory appears to reflect ‘recollection’ as opposed to ‘familiarity’ judgments.
2. Neurophysiology Of Visual Recognition
Although the neuronal mechanisms mediating visual recognition memory are not known, there is some information about the way in which neurons signal that a visual stimulus has been seen previously (Brown 1996). As shown in Figure 1, the temporal lobe consists of several parts. The MTL has already been discussed. On the outer, lateral surface of the temporal lobe are located visual areas TE and TEO. These regions are called the inferior temporal cortex (IT). The perirhinal and entorhinal cortex may be considered part of either IT or MTL. Many neurons in IT, including the perirhinal and entorhinal cortex, show changes in sensory responses with stimulus repetition. Two main kinds of recognition-related responses predominate.
Some neurons respond less to a visual stimulus on a monkey’s second or subsequent viewing of an object than they do on the first. Other neurons respond more. Similar phenomena occur in the medial thalamus and the prefrontal cortex, but not in the hippocampus. Thus, these types of activity signaling stimulus repetition occur in precisely those areas thought to be essential for performance on DNMS. In addition, there is a third type of neuronal activity related to recognition. In monkeys performing DMS (see Sect. 1.1), some neurons in both IT and the prefrontal cortex reflect information about the sample object during the interval between sample presentation and choice test, as though the cell were maintaining a memory of the sample (Desimone et al. 1995).
Furthermore, for prefrontal neurons, this type of activity persists even when other objects appear during a memory period. This kind of activity could be very useful for the short-term storage of information about an object and for directing attention to items of interest.
3. Cognition And Recognition
Recognition is only one among many kinds of memory and cognition. It means, literally, to ‘know again.’ In people, recognition can be normal, or nearly so, in the face of devastating memory impairments of other types—for example, an inability to remember personal daily activities. Conversely, recognition can be extremely impaired in people who are capable of learning and applying complex rules and strategies. Accordingly, recognition memory can be viewed as an aid to more complex cognitive operations, but is, itself, a modular function with adaptive advantages of its own. The nature of these advantages is uncertain, but probably involves the ability to direct attention to novelties in the environment (perhaps by reducing the sensory responses to recognized objects) or to familiar objects that are of importance to the behavior of the moment. It remains to be seen whether the mechanisms that underlie the recognition of objects, as discussed in this research paper, also subserve the recognition of thoughts and ideas, logical truth and falsity, esthetic beauty and ugliness, or other abstract aspects of human recognition that distinguish us from our fellow primates.
Bibliography:
- Aggleton J P, Brown M W 1999 Episodic memory, amnesia, and the hippocampal–anterior thalamic axis. Behavioral and Brain Science 22: 425–89
- Brown M W 1996 Neuronal responses and recognition memory. Seminars in Neuroscience 8: 23–32
- Desimone R, Miller E K, Chelazzi L, Lueschow A 1995 Multiple memory systems in the visual cortex. In: Gazzaniga M S (ed.) The Cognitive Neurosciences. The MIT Press, Cambridge, MA
- Mishkin M 1982 A memory system in the monkey. Philosophical Transactions: Royal Society London B 298: 85–95
- Mishkin M, Murray E A 1994 Stimulus recognition. Current Opinion in Neurobiology 4: 200–206
- Murray E A 2000 Memory for objects in nonhuman primates. In: Gazzaniga M S (ed.) The New Cognitive Neurosciences. The MIT Press, Cambridge, MA
- Parker A, Gaffan D 1998 Interaction of frontal and perirhinal cortices in visual object recognition memory in monkeys. European Journal of Neuroscience 10: 3044–57
- Squire L R 1987 Memory and Brain. Oxford University Press, New York
- Squire L R, Knowlton B J 2000 The medial temporal lobe, the hippocampus, and the memory systems of the brain. In: Gazzaniga M S (ed.) The New Cognitive Neurosciences. The MIT Press, Cambridge, MA
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality