
Sample Neural Basis Of Reading Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
Psychologists and brain researchers who investigate reading usually focus on the basic cognitive processes and brain regions that are necessary to recognize and pronounce a printed word. More complex aspects of reading, such as the ability to understand the grammatical roles played by each word in a written or spoken sentence, typically are assumed to rely upon general language functions that are not specific to reading. This research paper will focus on three components of skilled reading, and the brain regions that support them: a) the representation of visual word forms (orthography), b) the use of orthographic information to access information about word sounds (phonology), and c) the use of orthographic information to access information about meaning (semantics).
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
Claims about the neural bases of component processes associated with reading are based upon three primary methods. Studies of subjects with brain damage (neuropsychological studies) are used to understand how different aspects of reading performance are impaired and preserved following brain injury or degeneration. Studies of brain blood-flow changes (functional neuroimaging studies) use either PET (Positron emission tomography) or fMRI (functional magnetic resonance imaging) to localize and understand the functional contributions of brain regions associated with reading. ERP and MEG studies measure brain electromagnetic changes, and provide information about the temporal sequence of brain processing during reading, as well as some information about the location of brain regions associated with reading.
1. An Overview Of The Brain Regions Involved In Reading
An overview of the brain regions that support reading can be obtained by using neuroimaging to compare the pattern of brain activation associated with reading aloud a single word, versus merely looking at a small cross or dot. Some of the areas identified in this task comparison support relatively general sensory and motor functions. For instance, the primary motor cortex, which is located in the precentral gyrus (Fig. 1), is usually active during any task that incorporates movement. The identification of the motor cortex in the reading versus fixation comparison reflects the fact that a task that involves mouth movements is being compared to a task that does not. One approach for identifying regions that are more specifically involved in reading is to compare reading aloud to a control task that has similar sensory and motor components, but which minimizes orthographic, phonological, or semantic processing. For example, reading aloud has been compared to a task in which subjects say a simple utterance in response to a random string of letters. This type of contrast subtracts out basic visual, auditory, and motor areas, and reveals a more restricted set of regions that support higher-order aspects of reading (Fiez and Petersen 1998).
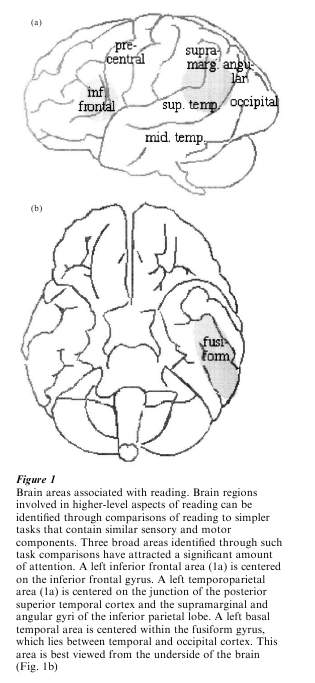
The exact number and location of the brain regions that support higher-order aspects of reading is a matter of active investigation, but three broad areas have attracted the most attention (Fiez and Petersen 1998; Price 1998). As illustrated in Fig. 1, these areas are centered in the left inferior frontal cortex, the left temporoparietal cortex, and the left basal temporal cortex (primarily the fusiform gyrus). Most of the remainder of this research paper will focus on the potential relationships between these brain areas and the basic components of skilled reading outlined in the introduction. It is important to note, as will become clearer in the discussion of the basal temporal area, that a more precise mapping of the brain regions that contribute to reading is still needed. It is likely that each of the broad areas focused on in this research paper contains multiple functionally distinct subregions. Furthermore, it is likely that other areas that are not discussed, such as the left middle temporal gyrus (Fig. 1), make some contribution to reading.
2. The Representation Of Orthography
The organization of the visual system has been a topic of longstanding interest in the neuroscience and cognitive neuroscience research communities. One question that is highly relevant for studies of reading is whether different regions of the visual object recognition system (located in the ventral occipital and temporal cortex) are specialized for processing different types of stimuli, such as faces or text. In the domain of reading, the search for a region that selectively processes text has generally involved two types of comparisons. The first has been comparisons of words and nameable pictures. This allows regions that are specifically involved in orthographic processing to be teased apart from regions that are more generally involved in the naming of meaningful stimuli. The second approach has been comparisons between different types of orthographic stimuli. Most often, words and pseudowords (pronounceable non-words, such as ‘floop’) have been compared to non-word stimuli that are visually similar to words (e.g., stimuli composed of artificial letters, or ‘false fonts,’ or random consonant strings). This allows regions that are specifically involved in the representation of words to be dissociated from regions that are involved in lower-level processes, such as letter recognition and the detection of line segments.
Several regions that may mediate orthographically specific processing have been proposed. The most promising ‘word form area’ is in the left middle fusiform gyrus, near the border between the temporal and occipital cortex. Activation in the fusiform gyrus is found reliably when subjects read words aloud (Fig. 1), and one reason to suspect that it may be involved in orthographic processing is the fact that it lies near areas associated with simple visual processing and visual object recognition. More direct evidence comes from studies that have found responses in the fusiform to words and pseudowords, but not visually similar nonword stimuli, such as random letter strings (Fiez and Petersen 1998). Evidence that this area is influenced by the degree to which an orthographic stimulus functions as a whole word (lexical) unit comes from studies in Japanese readers. The Japanese writing system contains two types of characters: Kana characters represent syllables, while Kanji characters represent entire words. Significant activation in the fusiform is found when subjects read words aloud written with Kanji versus Kana characters, and when subjects are given a task that requires them to access the orthographic form of a Kanji character, such as writing the Kanji equivalent of a word written in Kana (Nakamura et al. 2000, Sakurai et al. 2000).
Data from neuropsychology also point toward the important role of occipitotemporal areas, such as the fusiform gyrus, in orthographic processing. Subjects with brain damage in this area can exhibit the syndrome of pure alexia (Patterson and LambonRalph 1999). In pure alexia, the ability rapidly to access or represent the orthographic form of an entire word, or the ability to use orthographic information to access phonological and semantic information, is lost. As a result, subjects use a laborious letter-by-letter strategy to identify and read aloud a word. Subjects with pure alexia generally are able to perform simple visual perceptual tasks, recognize and name visual objects, and write words normally. Japanese subjects can also exhibit alexia following damage to the fusiform, but long-lasting impairments are typically limited to the reading and writing of Kanji characters (Iwata 1984).
Results from ERP and MEG studies indicate that there are differences in the electromagnetic current evoked by words and word-like stimuli, as compared to non-word stimuli such as letter strings (Cohen et al. 2000; Tarkiainen et al. 2000). The temporal information gained from these studies indicates that orthographically specific processing at the word level begins 100–200 milliseconds (ms) after the onset of a stimulus. Attempts to localize the source of the waveform differences have generally implicated the left basal temporal cortex. More specific localization has come from a study in which recordings were done using an electrode placed in the fusiform gyrus of patients prior to brain surgery. This study found neurons that selectively responded to orthographic stimuli (Nobre et al. 1994).
While there is a significant amount of accumulated evidence across multiple methodologies that points towards a role for the fusiform gyrus in orthographic processing, the precise function of this area remains a point of debate. One issue is whether domain-specific regions—‘face areas’ or ‘word form areas’—exist in the visual system. Since reading is a skill acquired very recently on the evolutionary time-scale, it is unlikely that any brain region would be innately specified to represent word forms. Rather, any specialization is most likely to occur as a result of extensive experience with a relatively unique class of visual stimuli. An alternative point of view is that the analysis of orthography places heavy demands on particular types of visual analysis, such as the ability to resolve input with high-frequency spatial information, or to represent a set of distinct objects that are perceptually very similar. Portions of the fusiform may be particularly suited for these types of analysis (either innately or through experience), and thus may be used to a greater extent for processing orthographic versus non-orthographic stimuli.
A second issue is the level of information that may be represented in the fusiform. Recent evidence indicates that letter-by-letter reading is typically accompanied by visual-perceptual difficulties below the level of the whole word. Deficits in lower-level processing could impair the access to or generation of higher-level orthographic representations (Behrmann et al. 1998). Conversely, imaging evidence reviewed more extensively below indicates that portions of the fusiform are active during linguistic tasks that are not thought to require access to orthographic information. Activation associated with orthographic processing may thus reflect more abstract lexical (whole word) or semantic information that can be accessed through either printed words or speech.
3. The Transformation Between Orthography And Phonology
In order to read a word aloud, it is essential that information about the printed form of the word is transformed into information about the corresponding sound, or pronunciation, of the word. Two general brain regions have been identified which appear to play a major role in phonological (word sound) representation and processing. One region is located in the left inferior frontal lobe, and the other is in the temporoparietal cortex.
Much of the neuropsychological evidence that these regions are important for reading stems from studies of patients with damage to either or both of these regions. These patients often exhibit the syndrome of phonological dyslexia, which is characterized by great difficulty reading aloud pseudowords (Fiez and Petersen 1998; Patterson and Lambon-Ralph 1999). The ability to read aloud pseudowords is considered to be a marker of phonological processing, because the pronunciation of pseudowords cannot be specified by stored lexical (whole word) or semantic knowledge. Instead, pronunciation is thought to rely upon some type of sublexical procedure for deriving a phonological representation. Subjects with phonological dyslexia can read real words relatively well. This is thought to reflect their ability to take advantage of stored lexical and semantic information using brain regions that are not damaged. Interestingly, when direct electrical stimulation is applied to the temporoparietal region during a reading task, subjects appear to exhibit a temporary phonological dyslexia: their ability to pronounce non-words is temporarily disrupted, but their reading of real words is intact (Simos et al. 2000).
Conclusions based upon neuropsychological data are supported by converging evidence from neuroimaging. Both the left frontal and temporoparietal regions are reliably active during normal word reading (Fig. 1), and during a wide variety of tasks that encourage phonological processing (Demonet et al. 1996). ERP and MEG evidence indicates that the activation of these areas occurs at least 300 ms after the onset of a stimulus, which is after the temporal differences observed in the basal temporal cortex (Simos et al. 2000). The importance of these regions in reading is further demonstrated by comparisons of normal adults to adults who experienced difficulties learning to read as children (developmental dyslexics). Many investigators believe that subjects with developmental dyslexia have a core deficit in phonological processing that persists into adulthood. When subjects with developmental dyslexia read words aloud, neuroimaging and MEG studies have revealed abnormally low levels of activity in the temporoparietal area (Shaywitz et al. 1998, Rumsey et al. 1999, Simos et al. 2000). The relative reductions in temporoparietal activation can be paralleled by above-normal levels of activity in the left frontal region. The left frontal increases may reflect use of a compensatory strategy for reading that normally plays a secondary role in pronunciation for skilled readers (except under more challenging conditions, such as when a stimulus is a nonword).
Several broad schemes that might distinguish the contributions of the frontal and temporoparietal regions to reading and phonological processing exist. One potential distinction is whether the phonological information and processes supported by a particular brain region are related to the acoustics of speech sounds (input phonology), or the articulation of speech sounds (output phonology). The proximity of the temporoparietal region to basic auditory areas, coupled with the fact that lesions to this area produce deficits in speech comprehension, indicates that it may support input phonology. Conversely, the proximity of the left frontal region to general motor-related areas, coupled with the fact that lesions to this area produce deficits in speech production, indicates that it may support output phonology.
A second potential distinction comes from broad differences between the roles of the frontal and posterior brain regions in cognition. Frontal regions are recruited most often by tasks that require conscious and effortful processing, or controlled processing. When a task is well learned and relatively automatic, activation in the frontal cortex is generally minimal. The frontal cortex may thus represent an area used for controlled phonological processing, whereas the temporoparietal region may represent an area used for automatic phonological processing. This could explain why subjects with developmental dyslexia, who do not develop the normal level of reading skills and automaticity, show less activation in the temporoparietal area and more in more frontal areas. It is also possible that different subregions within the inferior frontal cortex are responsible for different aspects of speech production (e.g., output phonology, grammatical analysis, etc.).
A third potential distinction comes from theoretical models of reading, which have outlined different types of mechanisms that could support orthographic to phonological transformation. Two broad classes of models have been proposed, dual-route (Coltheart et al. 1993) and connectionist models of reading (Plaut et al. 1996). In both types of models, access to the pronunciation of a word is mediated by two different ‘routes,’ though the routes function in different ways. In the dual-route models, a lexical route functions to associate orthographic information with stored representational information about the pronunciation of the whole word. A sublexical route functions to break a word or pseudoword into smaller parts, such as individual letters or letter clusters (graphemes), that can be associated with corresponding sounds (phonemes); the sounds can then be put together to yield a pronunciation for the entire item.
In connectionist models, direct mappings between orthography and phonology are used to capture statistical regularities between orthography and phonology at both the lexical (or whole word) and the sublexical (e.g., the ‘b’ in ‘back’ and ‘bear’ has the same sound) levels. The pronunciation of a written stimulus is represented as a distributed pattern of activity across a set of interconnected phonological units. Indirect mappings between orthography and phonology via semantics can also be used to support the pronunciation of a word. That is, you could represent the meaning of the word ‘dog,’ and also represent the fact that this meaning is associated with the pronunciation dog . As in the direct mappings between orthography and phonology, these mappings are represented as a distributed pattern of activity, but unlike the direct mappings the mappings should be relatively insensitive to sublexical information (since there is little relationship between parts of words and word meanings, at least for simple ‘mono-morphomic’ words like ‘dog’ and ‘table’).
Simple assumptions have been used to suggest potential mappings between theoretical components in models of reading and specific brain regions (Fiez and Petersen 1998, Price 1998). For instance, it has been suggested that the left frontal area contributes to a sublexical phonological process. This mapping is based upon the fact that in models of reading the pronunciation of pseudowords is primarily based upon output from a sublexical phonological procedure, and in the brain greater activation is seen in the left frontal area when subjects read pseudowords as compared to words. Even this potential correspondence, however, falls short of identifying whether the left frontal region functions according to dual-route or connectionist principles (or some alternative process), and it fails to acknowledge puzzling differences between words and pseudowords in other brain regions. In order to reach more complete levels of description, it will be essential to develop more rigorous methods for relating computational modeling and neuroimaging results.
4. The Transformation Between Orthography And Lexical Semantics
In addition to lexical procedures that facilitate direct associations between an orthographic and a phonological form of a word, it is also possible that other types of stored representational information can be used to support pronunciation. This could include lexical information about the word itself, the meaning of the word, or the concept denoted by the word. Once such lexical-semantic information has been accessed, it could be used to access information about the pronunciation of the word or concept, as suggested in connectionist models of reading. Importantly, access to most lexical-semantic information is expected to be modality and task independent. Either a written word or a picture, for instance, can be used to represent a word name or a semantic concept, and individuals can produce a spoken word in response to a written word, picture, or their own internal thoughts.
Of the three general areas that are commonly active during word reading (Fig. 1), the basal temporal region is the best candidate for an area that contributes to lexical-semantic processing. This is the same general region that has been implicated in orthographic representation, and the potential overlap between orthographic and lexical-semantic processing is discussed at the end of this section. Assertions that the basal temporal region contributes to lexical-semantic processing are based upon evidence that the region responds to a variety of stimuli and task conditions that emphasize the retrieval of word names or concepts. For instance, neuroimaging studies have found overlapping activation during picture naming and word naming tasks (Moore and Price 1999). Outside the domain of reading, basal temporal activation is commonly associated with tasks that emphasize semantic processing or word retrieval, such as deciding whether a pair of words belongs to the same semantic category (Price 1998).
Neuropsychological studies also point towards the role of the inferior temporal area in lexical-semantic processing. With respect to reading, the most critical information comes from studies of patients with acquired surface dyslexia. In the purest forms of surface dyslexia, subjects tend to mispronounce inconsistent words, which are words with unusual spelling-to-sound correspondences. For instance, the word ‘pint’ is not pronounced like ‘mint,’ ‘lint,’ or ‘hint.’ Theoretical models of reading have postulated that the pronunciation of inconsistent words, especially those that occur infrequently in print, is heavily dependent upon access to stored lexical semantic information. Most reported subjects with surface dyslexia have large regions of damage or degeneration affecting the left middle and inferior temporal lobe, and the deficits of these subjects typically extend beyond the domain of reading (Patterson and Lambon-Ralph 1999). For instance, they often have difficulty accurately naming pictures of objects. Surface dyslexics are generally able to read aloud most words with consistent spelling-to-sound correspondences, and pseudowords. This preserved ability may reflect the use of sublexical orthographic to phonological procedures that are supported by intact brain regions, such as the left frontal area.
An issue that is not well understood is whether the reading-related area discussed in Sect. 2 is the same as the lexical-semantic area discussed in this section. One reason to believe that they may be the same is the fact that in many neuroimaging studies of reading, only a single region of activation is identified in the fusiform gyrus. If they do overlap, future research will need to reconcile the orthographic and lexical-semantic points of view to provide a unified interpretation of the role of the inferior temporal area in reading. However, there is some evidence that there may be more than one functional region within the basal temporal area. A neuroimaging study of word versus picture naming, for example, identified multiple regions within the fusiform that had different patterns of activation (Moore and Price 1999). This is consistent with converging data from animal and human studies of visual object recognition. Such studies indicate that modality and domain specific visual processing, such as that involved in orthographic representation of lexical information, may be located in the posterior fusiform. In contrast, the anterior portions of the fusiform may be associated with increasingly conceptual and modality-independent processing. This could include the conceptual representation of objects and ideas that are denoted by pictures, and by written and spoken words (Ricci et al. 1999).
5. The ‘Tuning’ Of Brain Regions That Support Reading
Neuropsychological and neuroimaging studies of Chinese, Japanese, Italian, German, and English-speaking subjects have reached similar conclusions about the basic cognitive processes and brain regions involved in reading. This consistency reinforces the conclusion that the network of brain areas summarized in Fig. 1 is somehow specialized to support reading and other language processes. However, it should not be taken as evidence that the neural network that underlies reading is inflexible. Quite the contrary, there is growing evidence that a number of factors can have a profound impact on how the network is tuned to most effectively support reading.
Some of the tuning of the reading system may reflect differences in experience or training. For instance, in Italian the mapping between how words look and how they sound is much more consistent than it is in English. Italian-speaking subjects take advantage of this consistency to read aloud words and pseudowords more quickly than English-speaking subjects. Furthermore, when they read words aloud, they produce more brain activation in the left temporoparietal region than do English-speaking subjects; the converse is true in the left inferior temporal region (Paulesu et al. 2000).
Other differences may reflect the use of compensatory or alternative strategies. For instance, as noted above, subjects with developmental dyslexia show a hyperactivation of the left frontal region, and a hypoactivation of the left temporoparietal region. More subtle differences in the activation of brain areas associated with phonology are even apparent in readers at different ends of the normal distribution of reading skill (Rumsey et al. 1999). In subjects with acquired brain damage, neuroimaging studies indicate that right hemisphere brain regions, typically similar in location to sites of damage in the left hemisphere, may begin to play a more substantial role in language tasks (Buckner et al. 1996).
Bibliography:
- Behrmann M, Plaut D C, Nelson J 1998 A literature review and new data supporting an interactive account of postlexical effects in letter-by-letter reading. Cognitive Neuropsychology 15: 7–52
- Buckner R L, Corbetta M, Schatz J, Raichle M E, Petersen S E 1996 Preserved speech abilities and compensation following prefrontal damage. Proceedings of the National Academy of Science USA 93: 1249–53
- Cohen L, Dehaene S, Naccache L, Lehericy S, DehaeneLamberz G, Henaff M A, Michel F 2000 The visual word form area. Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain 123: 291–307
- Coltheart M, Curtis B, Atkins P, Haller M 1993 Models of reading aloud: Dual-route and parallel-distributed-processing approaches. Psychological Review 100: 589–608
- Demonet J F, Fiez J A, Paulesu E, Petersen S E, Zatorre R 1996 PET studies of phonological processing: A critical reply to Poeppel. Brain and Language 55: 352–79
- Fiez J A, Petersen S E 1998 Neuroimaging studies of word reading. Proceedings of the National Academy of Science USA 95: 914–21
- Iwata M 1984 Kanji versus Kana: Neuropsychological correlates of the Japanese writing system. Trends in Neuroscience 7: 290–3
- Moore C J, Price C J 1999 Three distinct ventral occipitotemporal regions for reading and object naming. Neuroimage 10: 181–92
- Nakamura K, Honda M, Okada T 2000 Participation of the left posterior inferior temporal cortex in writing and mental recall of kanji orthography. A functional MRI study. Brain 123: 954–67
- Nobre A C, Allison T, McCarthy G 1994 Word recognition in the human inferior temporal lobe. Nature 372: 260–3
- Patterson K, Lambon-Ralph M A 1999 Selective disorders of reading? Current Opinion in Neurobiology 9: 235–9
- Paulesu E, McCrory E, Fazio F, Menoncello L, Brunswick N, Cappa S F, Cotelli M, Cossu G, Corte F, Lorusso M, Pesenti S, Gallagher A, Perani D, Price C, Frith C D, Frith U 2000 A cultural effect on brain function. Nature Neuroscience 3: 91–6
- Plaut D C, McClelland J F, Seidenberg M S, Patterson K 1996 Understanding normal and impaired word reading: Computational principles in quasi-regular domains. Psychological Review 103: 56–115
- Price C J 1998 The functional anatomy of word comprehension and production. Trends in Cognitive Sciences 2: 281–8
- Ricci P T, Zelkowicz B J, Nebes R D, Meltzer C C, Mintun M A, Becker J T 1999 Functional neuroanatomy of semantic memory: Recognition of semantic associates. Neuroimage 9: 88–96
- Rumsey J M, Horwitz B, Donohue B C, Nace K L, Maisog J M, Andreason P 1999 A functional lesion in developmental dyslexia: Left angular gyral blood flow predicts severity. Brain and Language 70: 187–204
- Sakurai Y, Momose T, Iwata M, Sudo Y, Ohtomo K, Kanazawa I 2000 Different cortical activity in reading of Kanji words, Kana words and Kana nonwords. Cognitive Brain Research 9: 111–15
- Shaywitz S E, Shaywitz B A, Pugh K R, Fulbright R K, Constable R T, Mencl W E, Shankweiler D P, Liberman A M, Skudlarski P, Fletcher J M, Katz L, Marchione K E, Lacadie C, Gatennby C, Gore J C 1998 Functional disruption in the organization of the brain for reading in dyslexia. Proceedings of the National Academy of Sciences USA 95: 2636–41
- Simos P G, Papanicolaou A C, Breier J I, Fletcher J M, Wheless J W, Maggio W W, Gormley W, Constantinou J E C, Kramer L 2000 Insights into brain function and neural plasticity using magnetic source imaging. Journal of Clinical Neurophysiology 17: 143–62
- Tarkiainen A, Helenius P, Hansen P C, Cornelissen P L, Salmelin R 2000 Dynamics of letter string perception in the human occipitotemporal cortex. Brain 122: 2119–213
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality