
Sample Perceptual Organization Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
Unless we adopt a rather artificial analytical attitude, we perceive an environment neatly partitioned into objects rather than a mere collection of sensations. Here is a (red) cherry lying on a (yellow) plate, not merely a blob of red within a yellow surround. The concept of perceptual organization, the grouping of parts into larger units, originated with Gestalt psychology in its attack on the atomistic assumption of structuralism around the beginning of the twentieth century. A then widely held view was that complex percepts are simply additive combinations of sensory elements. Each sensory ‘atom’ was thought to be independent of all other atoms, tied together to larger complexes just by mechanisms of associative learning. The Gestaltists, on the contrary, emphasized that what we perceive is not ‘atomistic’ elements, but structured forms and shapes which interact within a perceptual ‘field’ of dynamic ‘forces.’ Perception was considered to rely on interactive processes within the nervous system determined by dynamic field forces.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Grouping Factors In Vision
Rules for how parts of visual stimuli are grouped together, ‘Gestalt laws,’ were first formulated by Max Wertheimer (1923) who essentially identified four factors: nearness (proximity), similarity, common fate, and closure.

1.1 Nearness Or Proximity
Other factors being equal, the nearer objects are to one another, the more likely they are to be organized into unified percepts. An example is shown in Fig. 1a, where the dots are easily organized into pairs of dots on the basis that each two neighboring dots form a unit. It is almost impossible to see the spaced dots as belonging together or, in other words, to break up the units based on nearness.
1.2 Similarity
The greater the similarity among objects, the more likely they are to be organized into perceptual units. For example, dots of the same color are readily seen as a group in distinction from dots of another color (Fig. 1b).
1.3 Common Fate
Dots which move simultaneously at the same rate and in the same direction, share a common fate, are readily seen as a group. The factor of common fate belongs to the strongest grouping factors; it ties together objects that are quite distant and different in form, size or color, given that they share a sort of event similarity in their sameness of motion. Where grouping would not otherwise occur in stationary displays, it will occur as soon as parts of the display move. A well-camouflaged animal will remain hidden only as long as it is standing still; it becomes visible (‘pops out’) as soon as it moves.
1.4 Closure
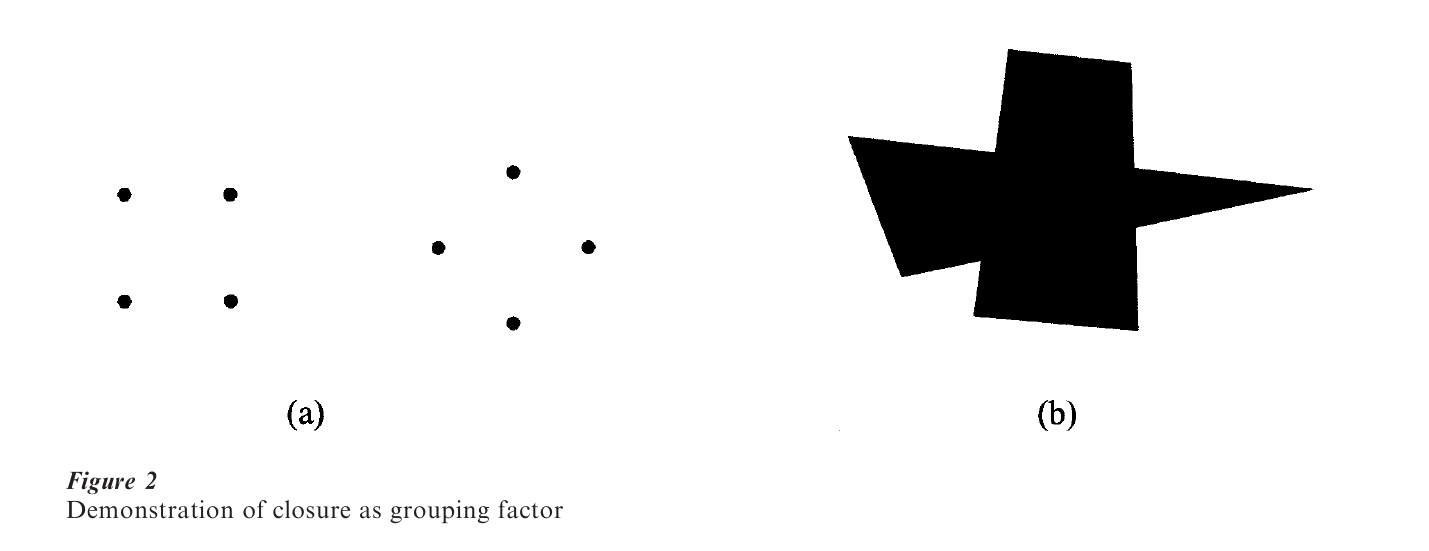
Components that constitute a closed entity rather than an open one are more readily organized into a single percept. This factor of closure predicts that a closed line has an advantage over an open one. For example, the two arrangements of four dots in Fig. 2a are seen as a rectangle or diamond rather than as crosses ( × ) or ( + ), because the former are closed. Closure also applies to the tendency to complete otherwise incomplete, e.g., partly occluded, objects. The configuration shown in Fig. 2b appears as a trapezoid overlapping a triangle rather than as a complicated eleven-sided figure.
1.5 Subjective Factors
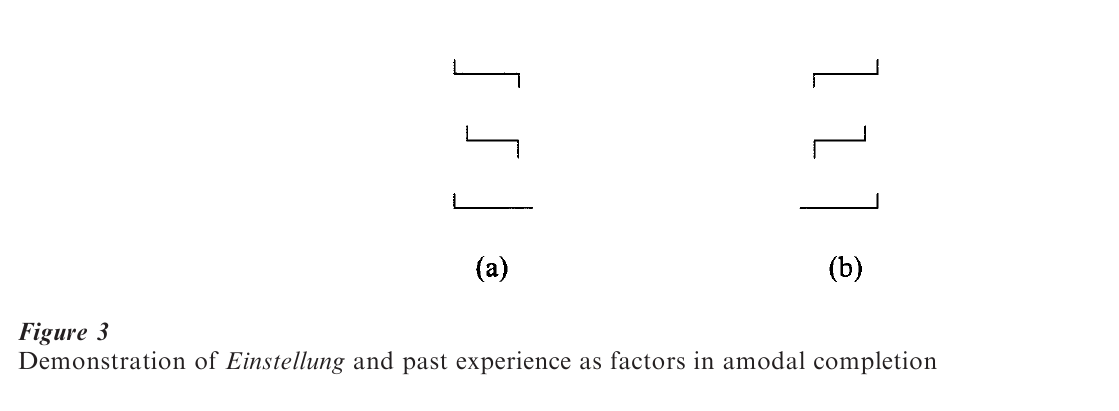
The study of natural camouflage and concealment shows that those grouping factors that refer to objective or stimulus variables fare well in accounting for the utility of various surface markings of animals. In addition, there are subjective or central factors at work, such as an observer’s momentary set, or Einstellung, and past experience, or custom, in determining how the perceptual field is organized. For example, the three separate lines shown in Fig. 3a may appear, at first glance, as a meaningless geometric arrangement of lines. However, the same configuration, just rotated by 180 (Fig. 3b), will be easily recognized as a capital E with the missing contours being perceptually completed. Now, if the reader with the Einstellung of an ‘E’ looks back at Fig. 3a, he or she will see a rotated E rather than a mere assembly of three lines. The perceptual formation of an E implies amodal completion of contours in the absence of stimulus (modal) information.
2. Figure–Ground Differentiation
Why do we see things rather than the holes between them? This question addresses another important aspect of organization, the differentiation of the perceptual field into figure and ground. This aspect was first brought out clearly by Edgar Rubin (1915). For example, if we look at a pencil on a desk, the pencil appears as a well-marked part of the perceptual field, the figure, while the desk appears as a relatively formless, featureless mass which we call the ground. The figure has the character of a thing, whereas the ground appears like unformed material. The ground seems to extend continuously behind or, sometimes, in front of the figure, i.e., although the figure usually appears in front, the ground is not always identical with background. For example, in looking through a window at a tree, the window screen appears as ground, but is clearly seen in front of the figure, the tree.
2.1 Dynamics Of Figure–Ground Perception
Rubin pointed out that even if all parts of a configuration are grouped together properly, it can appear either as object (figure) or surface (ground) under certain conditions. These so-called ambiguous or reversible figures ‘flip’ when, for example, the attention is directed to a specific point on the stimulus pattern. The dynamics of figure–ground perception due to the multivalence of the stimulus field are impressively demonstrated in a pattern shown in Fig. 4 (from Spillmann and Ehrenstein 1996)

New ‘flowers’ (figures) rise from the ‘meadow’ (ground) and disappear again, illustrating the vain attempt to establish a tendency toward a better and more stable percept. Apparently, the pattern allows for several groupings of equal probability and hence perception fails ever to reach a steady state. This example nicely illustrates that the visual space, as Kanizsa (1979, p. 181) artfully put it, is not a static geometrical scheme, nor is it a simple transposition on the perceptual level of the topographical arrangement of the retinal stimulus, but rather should be considered the result of an extremely dynamic event.
2.2 Tendency Towards Pragnanz
The Gestaltists assumed that the process of figural segregation and differentiation would follow a general tendency towards figural goodness or Pragnanz (Wertheimer), which denotes a processing principle for percepts to become structured in the most regular, symmetrical, and simple manner possible under the prevailing stimulus conditions. This process was understood by Wolfgang Kohler as a kind of self-organization according to electrodynamic field forces inherent in the sensory system. ‘In all processes which terminate in time-independent states the distribution shifts towards a minimum energy’ (Kohler 1920, quoted from Koffka 1935, p. 108). More than any other principle, the tendency towards Pragnanz emphasizes the global aspects of perceptual organization, since even minor local changes in the stimulus or in the observing process (such as eye movements or changes in attention) can result in a reorganization of the entire percept (see Fig. 4). A modern approach to the problem of self-organization and Pragnanz can be found in the theory of neural synergetics with its tendency towards ordered steady states in dynamic vector fields (Haken and Stadler 1990).
3. Perceptual Organization In Other Sense Modalities
Although the principles of perceptual organization have been studied most thoroughly for the visual modality, they also apply to other sense modalities. For auditory perception, Bregman (1990) has provided many examples demonstrating the resemblance between audition and vision. Temporal proximity of tones is more important than spatial proximity: Tones that follow each other close together in time are perceived as belonging together. Similarity is determined primarily by the pitch of tones: tones similar in pitch tend to be grouped together in perception. When a sequence of alternating high and low-frequency tones is played at a certain rate, listeners are able to follow the entire sequence of tone, provided that the frequency difference between tones is small and the tones are played at a slow rate. With greater frequency differences or higher rates, however, the sequence splits into two streams, one high and one low in pitch (auditory stream segregation).
Other examples—studies in tactile perception, for instance—confirm the notion that the above-mentioned factors of perceptual grouping play a similar role in different modalities. This suggests that the perceptual system can utilize organizing principles from one sensory modality to the other, i.e., in a cross-modal way. Moreover, cross-modal interactions can change the percept. A recent example concerns the perception of emotions. Listeners having to judge the emotion in the voice are influenced by whether the (visually displayed) face expresses the same emotion or a different one (De Gelder and Vroomen 2000).
4. Neural Correlates Of Perceptual Organization
More than 70 years after Wertheimer’s pioneering formulation of perceptual grouping factors, terms that were originally introduced to describe qualitatively properties of figural grouping and segregation are being used as a much-needed vocabulary for interpreting spatiotemporal processing of sensory neurons and neuronal systems. The Gestaltists themselves, notably Wolfgang Kohler, tried to link perceptual organization to brain processes. Inspired by the laws of thermodynamics, the brain was thought to be a homogeneous conductor of bioelectric forces rather than a complex network of interacting neurons (e.g., Kohler and Held 1949). However, the field forces postulated by Kohler as factors in perceptual organization could not be substantiated by physiological evidence. Neural correlates of perceptual grouping are today assumed to consist of receptive field organization and synchronized coupling of neuronal activity within and across cortical areas (e.g., Spillmann and Ehrenstein 1996).
4.1 Receptive Field Organization: A Micro-Gestalt
The processing of sensory input obeys essentially two principles, convergence and divergence. Many receptors send their input to the same ganglion cell (convergence) and conversely one and the same receptor distributes signals to several ganglion cells (divergence). The receptor surface area within which a change of stimulation can modify the firing rate of a given neuron is called its receptive field. In vision, receptive fields of retinal, geniculate, and cortical neurons are concentrically organized with an excitatory center and an inhibitory surround (on-center neurons) or with a center that is inhibited by light and a surround that is activated by light (off-center neurons). The functional unit represented by the receptive field (with its center-surround antagonism and selective spatial and temporal sampling) is a basic neuronal mechanism that achieves non-additive integration of stimulus input. It has therefore been considered a micro-Gestalt by Spillmann and Ehrenstein (1996) with the conjecture that perceptual organization is largely determined by receptive-field organization.
4.2 A Neurocomputational Example
A detailed neurocomputational model of how perceptual groupings might emerge from interactions of cells with known receptive-field properties of the lateral geniculate and cortical areas V1 and V2 has been recently proposed by Grossberg et al. (1997). For example, global properties of Gestalt grouping (as demonstrated in Fig. 5) can be accounted for by their model that involves multiple levels of thalamo-cortical interaction to generate emergent boundary groupings.
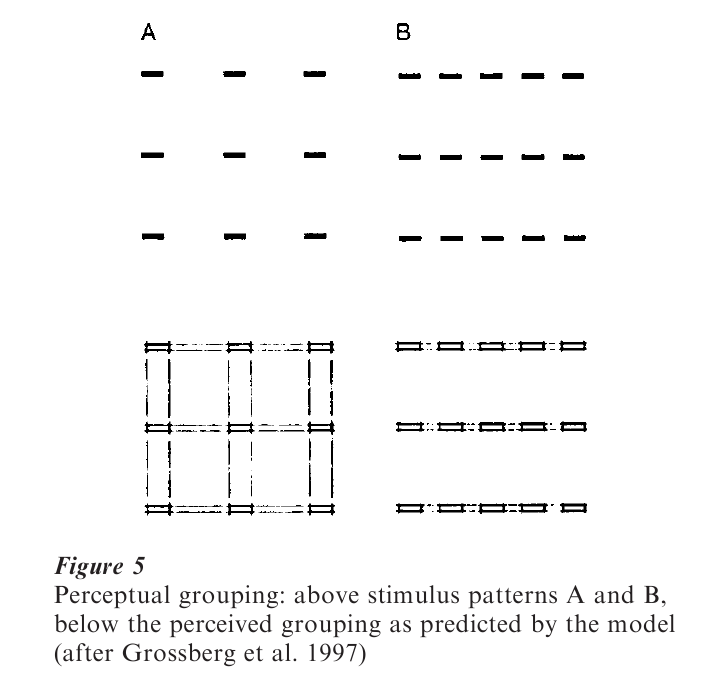
The left side of Fig. 5 (from Grossberg et al. 1997) shows an ambiguous configuration (A, upper part) which allows the perception of vertical or horizontal groupings of bar elements and the simulated outcome by the neurocomputational model (A, lower part). Adding horizontal bar elements (Fig. 5, B) leads to horizontal grouping in perception (upper part) and also in the model (lower part).
4.3 ‘Perceptive’ Neurons And Synchronized Coupling Of Spike Activity
In a few cases ‘perceptive’ neurons (Baumgartner 1990) have been reported that mediate invariant stimulus properties, e.g., neurons in area V4 of primate cortex seem to signal the perceived color of a stimulus, irrespective of its spectral composition and changes in illumination. The striking correlations between the activity of highly specialized single neurons and complex perceptual performance support the view that neurons at higher levels in the processing hierarchy may be sufficient to convey Gestalt perception. However, search for individual neurons responding selectively to complex configurations has, so far, revealed a very limited set of biologically significant stimuli. Alternatively, synchronized oscillations of even far-distant neurons allow for perceptual coherence over a large area of the sensory field. The above-mentioned grouping factor of common fate could especially be accounted for by temporal binding of features (e.g., Eckhorn 1999). In fact, synchronized coupling of spike activity between remote brain areas has been found to be context-dependent and to account for spatiotemporal coherence in perception. Moreover, synchronized feature binding affords individual cells and cell groups flexibly to change partners with which they share activity when the stimulus configuration changes. Thus, single-cell activity, organized within a hierarchy of receptive fields, and synchronized feature binding seem to provide the basic neural mechanisms subserving perceptual grouping.
Bibliography:
- Baumgartner G 1990 Where do visual signals become a perception. In: Eccles J C, Creutzfeldt O (eds.) The Principles of Design and Operation of the Brain. Springer, Berlin, pp. 99–114
- Bregman A S 1990 Auditory Scene Analysis. MIT Press, Cambridge, MA
- De Gelder B, Vroomen J 2000 The perception of emotion by ear and eye. Cognition and Emotion 14: 289–311
- Eckhorn R 1999 Neural mechanisms of visual feature binding investigated with microelectrodes and models. Visual Cognition 6: 231–65
- Grossberg S, Mingolla E, Ross W D 1997 Visual brain and visual perception: How does the cortex do perceptual grouping? Trends in Neurosciences 20: 106–11
- Haken M, Stadler M (eds.) 1990 Synergetics of Cognition. Springer, Berlin
- Kanizsa G 1979 Organization in Vision. Essays on Gestalt Perception. Praeger, New York
- Koffka K 1935 Principles of Gestalt Psychology. Harcourt Brace, New York
- Kohler W, Held R 1949 The cortical correlate of pattern vision. Science 110: 414–19
- Rubin E 1915 Synsoplevede Figurer. Gyldendal, Copenhagen, Denmark
- Spillmann L, Ehrenstein W H 1996 From neuron to Gestalt. Mechanisms of visual perception. In: Greger R, Windhorst U (eds.) Comprehensive Human Physiology. Springer, Berlin, Vol. 1, pp. 861–93
- Wertheimer M 1923 Untersuchungen zur Lehre von der Gestalt. II. Psychologische Forschung 4: 301–50
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality