
Sample Neural Basis Of Declarative Memory Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. iResearchNet offers academic assignment help for students all over the world: writing from scratch, editing, proofreading, problem solving, from essays to dissertations, from humanities to STEM. We offer full confidentiality, safe payment, originality, and money-back guarantee. Secure your academic success with our risk-free services.
Memory is a broad topic that has its roots in both biology and psychology. Many important aspects of the problem of memory involve systems-level questions that must be addressed at a relatively global, neuropsychological level of analysis. Two long-standing and inter-related questions have been of central interest since the 1960s. The first is whether there is more than one kind of memory. The second concerns what brain structures and pathways are involved in memory. Scientific work on these issues has introduced the term declarative memory to neuroscience and identified a number of parallels between the neural organization of memory in humans and animals.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
It has turned out that the brain is organized such that memory is a distinct and separate cognitive function, which can be studied in isolation from perception and other intellectual abilities. Significant information about how memory is organized has come from two experimental approaches. First, the study of patients with memory disorders (amnesia) has been enormously useful. Second, information about the anatomy of memory has come mainly from the kind of systematic and cumulative work that is possible only in experimental animals.
1. Amnesia And The Medial Temporal Lobe Memory System
Amnesia refers to difficulty in learning new information or in remembering the past. Amnesia can occur as a strikingly circumscribed memory impairment in the absence of other cognitive deficits (Scoville and Milner 1957). Patients with this disorder can have intact intelligence test scores, intact language and social skills, and an intact memory for a great deal of information from the remote past, especially child-hood. In addition, immediate memory is intact. Indeed, patients can hold information in mind for several minutes, provided there is no distraction and the material can be rehearsed. Their intact immediate memory explains why amnesic patients can carry on a conversation and can appear quite normal to casual observation. The difficulty for individuals with amnesia arises when information must be recalled after a distraction-filled interval, or when recall is attempted directly after the presentation of an amount of information that exceeds immediate memory capacity (e.g., a list of eight or more items). In this situation, normal individuals will usually recall more items than amnesic patients.
An animal model of human amnesia was established in the monkey in the early 1980s (Mishkin 1982, Squire and Zola-Morgan 1983). Systematic and cumulative work during the next 10 years, using the animal model, succeeded in identifying the system of structures in the medial temporal lobe essential for memory (Squire and Zola-Morgan 1991) (Fig. 1). This system includes the hippocampal region (i.e., the hippocampus proper, the dentate gyrus, the subicular complex), the entorhinal cortex, and the adjacent perirhinal and parahippocampal cortices. A number of studies in monkeys have been carried out, comparing performance on memory tasks for monkeys with damage to different components of the medial temporal lobe. This work has shown that the severity of memory impairment depends on the locus and extent of damage within the medial temporal lobe memory system. Damage limited to the hippocampal region causes significant memory impairment, but damage to the adjacent cortex in-creases the severity of memory impairment. It is important to note that the discovery that larger medial temporal lobe lesions produce more severe amnesia than smaller lesions is compatible with the idea that structures within the medial temporal lobe might make qualitatively different contributions to memory function. This is because anatomical projections carrying information from different parts of the neocortex enter the medial temporal lobe memory system at different points (Suzuki and Amaral 1994).
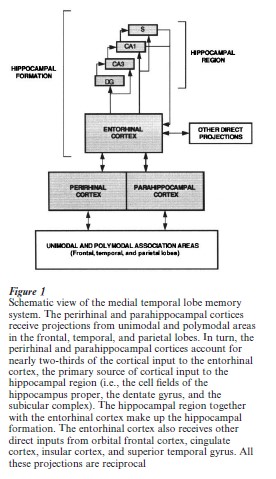
Another important brain area for memory is the diencephalon, However, the critical regions in the diencephalon that when damaged produce amnesia have not at the time of writing been identified with certainty. The important structures appear to include the mediodorsal thalamic nucleus, the anterior nucleus, the internal medullary lamina, the mammillo-thalamic tract, and the mammillary nuclei. Because diencephalic amnesia resembles medial temporal lobe amnesia in many ways, these two regions together probably form an anatomically linked, functional system.
At about the same time that an animal model of amnesia was established, detailed case studies of human amnesia provided extensive neuropsychological information about memory impairment, as well as detailed neuropathological information about the locus and extent of damage in each patient’s brain. New techniques for imaging brains have also made it possible to detect some kinds of neuropathology in living patients. The available neuroanatomical in-formation from humans is consistent with the findings from monkeys. These lines of work have greatly clarified the description of memory impairment and its anatomical basis, and have made it possible to pursue parallel work in simpler animals such as rats and mice. As a result, one can now study memory in rodents and have some confidence that what one learns can be relevant to the human condition.
2. Declarative Memory
An important finding of neuropsychological studies of memory is that memory is not a single entity but is composed of several separate systems (Squire 1992, Schacter and Tulving 1994). Figure 2 illustrates how long-term memory can be divided into several parallel memory systems. The major distinction is between conscious knowledge of facts and events, referred to as declarative or explicit memory, and other non-conscious knowledge systems that provide for the capacity of skill learning, habit formation, the phenomenon of priming, and certain other ways of interacting with the world, collectively referred to as nondeclarative or implicit memory. Nondeclarative memory is not itself a brain-systems construct. Rather, this term encompasses several different kinds of memory. Nondeclarative forms of memory have in common the feature that memory is nonconscious. Memory is expressed through performance and does not require reflection on the past or even the knowledge that memory is being influenced by past events.
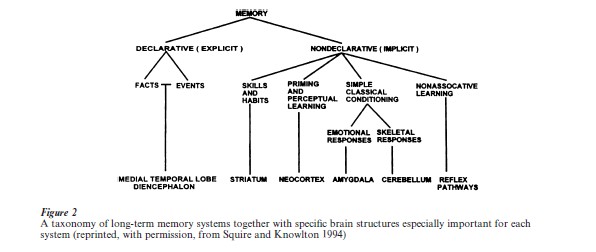
Declarative memory (explicit memory, relational memory) is a brain-systems construct, referring to the capacity for conscious recollections about facts and events that is dependent on the integrity of the hippocampus and anatomically related structures in the medial temporal lobe and diencephalon. It seems to be especially good at associating the various aspects of a context that are present at a particular time and place, thereby creating a memory of an episode. Declarative memory is also well suited for connecting the pieces of information needed to acquire a new fact (e.g., the capital of North Dakota is Bismarck). It is sometimes pointed out that declarative memory allows one to model the external world, and it is either true or false; nondeclarative memory is about performance, and it is neither true nor false.
Declarative memory is the kind of memory impaired in amnesia. However, the concept of declarative memory is not defined solely in terms of what amnesic patients can and cannot learn. Other characteristics of declarative memory have also been identified. For example, work with amnesic patients has emphasized the notion of conscious recollection, though this description is unhelpful when considering learning and memory in animals. Other characteristics have emerged from the studies of experimental animals. Declarative memory is fast, it is not always reliable (i.e., forgetting and retrieval failure can occur), and it is flexible in the sense that it is accessible to multiple response systems. It is especially suited for one-trial learning, and forming and maintaining an association between two arbitrarily different pieces of material.
2.1 Recognition Memory
A particularly good example of declarative memory involves the capacity to identify a recently encountered item as familiar, a capacity termed recognition memory. Recognition memory is impaired in amnesic patients, and it is impaired in monkeys and rats following damage to the hippocampal region. In the monkey, the assessment of recognition memory has depended mainly on two tasks. In delayed nonmatching to sample, two objects are presented, a new one and one that was presented earlier. The monkey must choose the new object. In the visual paired-comparison task, as adapted for the monkey, a new picture and a recently presented picture are presented side by side, and the monkey’s spontaneous tendency to look at the new picture is measured. When the monkey shows a preference for viewing the new object, the monkey is demonstrating that it remembers the old object.
At the time of writing (2000), a total of six studies from different laboratories have been carried out on the effects of lesions of the hippocampal region on memory in monkeys. The lesions were made by an ischemic procedure, by radiofrequency, or ibotenic acid. Postmortem neurohistological analysis of the lesions is available in all six studies. Five of the studies have found impaired recognition memory. The sixth study, which differed from the others in potentially important ways, found no impairment (see Zola et al. 2000 for a review of these studies).
The evidence that the hippocampal region is essential for normal recognition memory is entirely consistent with current ideas about the role of the hippocampus in declarative memory and with the view that the hippocampus is essential for acquiring information about relationships, combinations, and conjunctions among and between stimuli (Eichenbaum 1997). The recognition test asks whether an item that had been presented recently now appears familiar. A recognition (or familiarity) decision re- quires that the stimulus presented in the retention test be identified as what was presented during learning. At the time of learning a link must therefore be made between the to-be-remembered stimulus and its con- text or between the stimulus and the animal’s interaction with it. It is this associating and the ability to retain relational information across time that many have supposed is at the heart of declarative memory and in turn is the function of the hippocampal region in both humans and nonhuman animals.
2.2 Episodic And Semantic Memory
Episodic memory refers to the capacity for recollecting an autobiographical memory of events that occurred in a particular spatial and temporal context. Semantic memory refers to the capacity for recollecting general knowledge and facts about the world. Both types of memory are declarative in the sense that subjects are retrieving information explicitly, and they are aware that stored information is being retrieved. There is good agreement that episodic memory is impaired in amnesia following damage to the medial temporal lobe memory system. However, there is less agreement about semantic memory and its relationship to the memory system. By some accounts, amnesic patients are proportionately impaired in both episodic and semantic memory. By other accounts, the capacity for semantic memory is spared, or partially spared in amnesia relative to episodic memory ability, especially if the damage is limited to the hippocampal region.
Evidence for the second view comes especially from two kinds of data. First, there has been interest in cases where amnesia occurred in early childhood, before much of an individual’s semantic knowledge has been acquired (Vargha-Khadem et al. 1997). Second, there have been studies of amnesic patients where the ability to accomplish fact learning and event learning has been directly compared. Taken together, the data tend to favor the view that episodic and semantic memory are similarly impaired in amnesia (Squire and Zola 1998). Other work suggests that episodic memory additionally depends on the frontal lobes, which provide a source of autobiographical involvement with the recollected material. This orientation allows for a fundamental distinction between episodic and semantic memory, but without depending on differences in how the two kinds of memory depend on medial temporal lobe structures.
3. Overview
Work with amnesic patients and experimental animals has converged to characterize the organization of declarative memory. Declarative memory depends on the integrity of the medial temporal lobe, which works in concert with the neocortex. Declarative memory may develop gradually from early in life. Human infants six months of age successfully perform the visual paired-comparison task, and performance on this task is impaired by hippocampal lesions in humans and monkeys. The view that some declarative memory is available early in life is also consistent with demonstrations of long-term, recall-like memory abilities in human infants.
Other work with amnesic patients has suggested that declarative memory is equivalently involved in the two fundamental processes of recall and recognition. Work in humans and animals has shown that the medial temporal lobe memory system is required at the time of learning if an enduring and retrievable long-term declarative memory is to be established. Studies of retrograde amnesia in amnesic patients and in experimental animals suggest that the brain system supporting declarative memory has a time-limited function. Medial temporal lobe lesions spare very remote memories. Thus, as time passes after learning, a process of consolidation and reorganization occurs such that permanent storage does not require the medial temporal lobe. Permanent memory is thought to be stored in the neocortex where information is received and processed. Information is still accumulating about how memory is organized, what structures and connections are involved, and what functions they support. The disciplines of both psychology and neuroscience continue to contribute to this enterprise.
Bibliography:
- Eichenbaum H 1997 Declarative memory: insights from cognitive neurobiology. Annual Review of Psychology 48: 547–72
- Mishkin M 1982 A memory system in the monkey. Philosophical Transactions of the Royal Society London—Biological Sciences 298: 85–92
- Schacter D 1996 Searching for Memory. Basic Books, New York
- Schacter D L, Tulving E (eds.) 1994 Memory Systems. MIT Press, Cambridge, MA
- Scoville W B, Milner B 1957 Loss of recent memory after bilateral hippocampal lesions. Journal of Neurology, Neurosurgery, Psychology 20: 11–21
- Squire L R 1992 Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychological Review 99: 195–231
- Squire L R, Kandel E R 1999 Memory: From Mind to Molecules. Freeman, New York
- Squire L R, Zola S M 1998 Episodic memory, semantic memory, and the medial temporal lobe memory system. Hippocampus 8: 205–11
- Squire L R, Zola-Morgan S 1983 The neurology of memory: The case for correspondence between the findings for human and nonhuman primate. In: Deutsch J A (ed.) The Physiological Basis of Memory. Academic Press, New York
- Squire L R, Zola-Morgan S 1991 The medial temporal lobe memory system. Science 253: 1380–6
- Suzuki W A, Amaral D G 1994 Perirhinal and parahippocampal cortices of the macaque monkey: cortical aff Journal of Computational Neurology 350: 497–533
- Tulving E, Craik F I M (eds.) 2000 The Oxford Handbook of Memory. Oxford Press, New York
- Vargha-Khadem F, Gadian D G, Watkins K E, Connely A, Van Paesschen W, Mishkin M 1997 Differential effects of early hippocampal pathology on episodic and semantic memory. Science 277: 376–80
- Zola S M, Squire L R, Teng E, Stefanacci L, Clark R E 2000 Impaired recognition memory in monkeys after damage limited to the hippocampal region. Journal of Neuroscience 20: 451–63
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality