
Sample Neural Basis of Memory and Aging Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
A predominant view of the neural basis of memory and aging in the twentieth century was that tens of thousands of neurons were lost on a daily basis in older adults resulting in impaired cognition and memory in old age. The early twenty-first century view of this phenomenon has changed rather dramatically.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
In the 1990s there was growing recognition of the fact that neuronal numbers in many brain structures remain relatively constant. Nevertheless, on a functional level, learning and memory are impacted by processes of aging. Research on the neural basis of memory and aging has identified neurochemical and electrophysiological correlates of age-related memory impairment in some older animals, including some older humans. Other older individuals show little impairment. The inclusion of older adults in early stages of neurodegenerative diseases in studies of normal aging has also contributed to a perspective of functional decline in the whole population. Both the magnitude of neuronal loss and the degree of functional memory impairment were likely overestimated in previous research.
1. Neuronal Counts and Aging
It has become evident with advances in research techniques and technology that traditional cell counting methods were flawed, often under-representing the number of cells in older tissue. Stereological cell counting techniques that provide unbiased estimates of cell numbers have replaced cell density measures. The application of stereological techniques in studies of aging in the nervous system has resulted in a revision of the view that large numbers of neurons die during normal aging, at least for some structures. Agerelated loss of larger numbers of neurons in memoryrelated circuits does not appear to be a concomitant of normal aging but rather a consequence of dementing diseases such as Alzheimer’s disease (AD; Morrison and Hof 1997). Medial-temporal lobe structures including the hippocampus and the neocortex are essential for the form of memory used for conscious recollection of facts and events. Although there is agerelated decline in this form of memory, neuronal loss in the medial-temporal lobes does not appear to be the cause of functional impairment.
2. Brain Memory Systems, Normal Aging, and Neuropathological Aging
Human long-term memory systems have been classified into two major forms: declarative (or explicit) and nondeclarative (implicit or procedural). Declarative learning and memory, which can be conceptualized as learning with awareness, refers to the acquisition and retention of information about events and facts. It is assessed by accuracy on recall and recognition tests (e.g., word-list learning). Four decades of research support the conclusion that medial-temporal diencephalic memory circuits are critical for establishing long-term memory for events and facts (Squire 1992). In contrast, there is considerable evidence that medial-temporal diencephalic memory circuits are not critical for several kinds of nondeclarative memory.
Brain substrates of declarative and nondeclarative memory systems can function in parallel and independently from one another. Within the nondeclarative category, brain substrates of various forms of learning and memory are often physically remote from one another and composed of qualitatively different types of neurons, neurotransmitters, and projections. Most importantly for an understanding of the neural basis of memory and aging, the various brain memory systems are probably affected differentially by processes of aging.
2.1 Declarative Learning and Memory
Among brain memory systems, it is the declarative form that draws the greatest research attention in gerontology. The subjective awareness of ‘trying to learn’ is typically present in tasks that assess declarative learning and memory. Lesions in the medialtemporal lobe region including the hippocampus are associated with deficits in declarative learning and memory. It has become increasingly evident that lesions in these regions are not the consequence of normal aging but rather a part of the neuropathology of AD. Early in the course of AD, impairments in memory and cognition are hardly detectable. It is difficult to distinguish these patients with early AD from normal older adults. Yet, in the earliest stages of the disease, as many as half of the neurons in some critical layers of the medial-temporal lobes essential in the declarative memory circuit are lost. Without a doubt, many studies of normal aging have included in the sample individuals with early AD.
For example, there was evidence of hippocampal atrophy from radiological brain scans in about onethird of 154 community-residing older adults ranging in age from 55 to 88 years. These ‘normal’ older adults participated in a longitudinal study of aging and memory (Golomb et al. 1993). It was primarily the participants with hippocampal atrophy who performed poorly on tests of immediate and delayed recall of prose paragraphs and paired associate words. Participants with hippocampal atrophy were probably in the early stages of AD. Stereological cell counting techniques of post mortem brain tissue demonstrated that almost complete loss of pyramidal cells in the CA1 field of the hippocampus was a characteristic feature of AD that did not occur in normal aging (West et al. 1994).
Magnetic resonance imaging (MRI) assessment of hippocampal atrophy has become useful in the diagnosis of AD early in the course of the disease (Jack et al. 1997). Older adults with mild cognitive impairment who are at risk of AD have significant volume reduction in the hippocampus. Furthermore, individuals at risk of developing AD but presymptomatic at the initial testing develop memory impairment that is paralleled with hippocampal loss when followed up a year or two after initial MRI testing. There are observations from many laboratories including our own of an association between hippocampal atrophy in the early states of degenerative disease and declarative memory impairment (see Fig. 1).
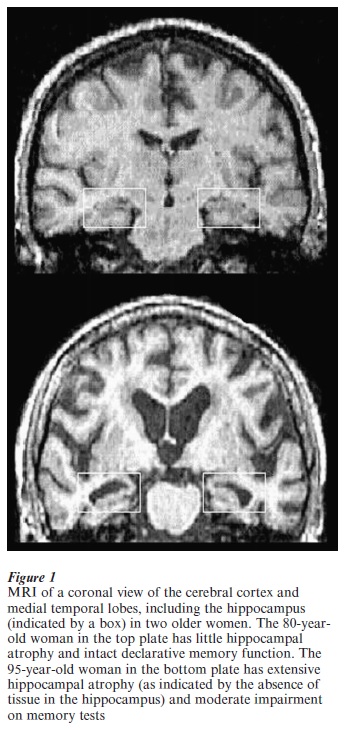
Whereas there is only limited neuronal loss in medial-temporal lobe regions in normal aging, declarative memory shows functional age-related deficits in a majority of older adults, with recall affected more than recognition. Age differences in memory processes such as the encoding of new information that engages the prefrontal cortex as well as medial-temporal lobes have also been documented. If neuronal loss cannot account for the cognitive impairment, what neural mechanisms might cause the behavioral deficits?
Age-related changes in neurotransmitter levels and in electrophysiological properties have been associated with impaired memory. However, these neurobiological changes vary among individuals. Individual differences in memory abilities are larger in old age in a number of species, with some older animals performing as well as the young, while other older animals are seriously impaired. At a neurotransmitter level, there is evidence that the wide variation in acetylcholine input to the hippocampus is associated with the greater variability in memory ability in older rats. A marker for acetylcholine, choline acetyltransferase (ChAT), was significantly reduced in the medial septal region only in older rats with memory impairments (Gallagher and Rapp 1997). Nevertheless, the ChAT levels and memory performance in some older rats is equal to that of young rats. There are many electrophysiological properties of neurons in aging rodents that remain intact, but one learning and memoryrelated phenomenon that may become impaired is long-term potentiation (LTP; Barnes 1998). Again, the pattern is that some older organisms that exhibit severely impaired performance on memory tasks have lower levels of LTP, but some older organisms have LTP and memory performance levels similar to levels in young animals.
2.2 Nondeclarative Learning and Memory
Nondeclarative learning and memory may be conceptualized as ‘learning without awareness’ that is measured by changes in performance. Nondeclarative learning and memory consist of multiple, dissociable processes, including (a) repetition priming effects, (b) the acquisition and retention of motor, perceptual, or problem solving skills, and (c) simple forms of classical conditioning. Repetition priming is assessed by a decrease in reaction time or a bias in response to particular words or patterns as a consequence of prior exposure (during the experiment) to those words or patterns. Skill learning is assessed by improvement in speed and accuracy across trials on repetitive sensorimotor tasks. Classical conditioning is assessed with the pairing of a neutral and a reflex-eliciting stimulus and is observed when the neutral stimulus elicits the reflexive response. The various neural substrates of nondeclarative learning and memory are physically remote from one another and have little overlap. Brain imaging studies in humans have demonstrated that circuitry in the cerebellum is activated in eyeblink classical conditioning. Motor skill learning engages the motor cortex. Repetition priming involves the occipital cortex with the frontal cortex engaged in priming tasks requiring production.
The initial brain memory systems perspective of aging was that nondeclarative forms of learning and memory remained relatively intact whereas declarative forms were impaired. In the case of repetition priming, additional investigation has revealed a more complex perspective. An overview of the literature indicates a mild but consistent reduction in priming in normal aging, but the underlying substrate for this result is unknown. Mild age-related impairment in repetition priming may reflect reduced priming in a subgroup of older adults who are aging less successfully or have as yet undetected AD, or it may represent an impairment that all older adults experience as a consequence of normal aging (Fleischman and Gabrieli 1998).
The studies that have examined motor skill learning in normal aging are not consistent in their results, but the perspective again is one of minimal age-related effects. Various studies of skill learning and aging have reported three different kinds of results: superior performance by older adults, equal performance by young and old, and superior performance by young adults. Comparing age-matched older adults to patients with a diagnosis of probable AD, Eslinger and Damasio (1986) found no differences in acquisition of a rotary pursuit task. However, the AD patients were seriously impaired on tests of declarative memory. Again, the perspective for motor skill learning is that there is a mild effect of normal aging that is less dramatic than observed on declarative memory tasks.
A different perspective emerges from studies of simple eyeblink classical conditioning. Since the first studies were carried out in the 1950s comparing young and older adults on this task, striking age differences were apparent. When adults over the age range of 20 to 90 are tested, age-related effects appear in the decade of the 40s. Direct comparisons of a declarative memory measure (California Test of Verbal Learning) and a nondeclarative measure (400 ms delay eyeblink classical conditioning) in the same young and older adults revealed a larger age effect on the nondeclarative than the declarative task (Woodruff-Pak and Finkbiner 1995).
Like all investigations of normal aging, when participants have subclinical pathology (as in the early stages of AD) performance is affected on eyeblink classical conditioning. Indeed, several laboratories have demonstrated that patients diagnosed with probable AD are severely impaired on this task. However, in elderly adults with normal neuropsychological profiles, it is the status of the cerebellum that is related to performance on eyeblink conditioning. The correlation between eyeblink conditioning performance and cerebellar volume corrected for total brain volume was .81 ( p .02) in a sample ranging in age from 77 to 95 years (Woodruff-Pak et al. 2000).
Until quite recently, the role of the cerebellum in cognition received little research attention whereas the role of medial-temporal lobe structures including the hippocampus in memory has been a central focus of research in cognitive neuroscience. Studies of cognition in normal aging have overlooked the potential role of the cerebellum. There are striking differences in cerebellar volume between young and older adults (see Fig. 2), and tasks that are cerebellar-dependent show age-related deficits earlier than hippocampally-dependent tasks. Further exploration of the role of the cerebellum in cognitive and aging processes is warranted.
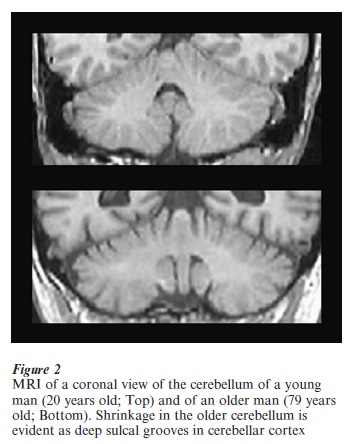
Bibliography:
- Barnes C A 1998 Spatial cognition and functional alterations of aged rat hippocampus. In: Wang E, Snyder D S (eds.) Handbook of the Aging Brain. Academic Press, San Diego, CA
- Eslinger P J, Damasio A R 1986 Preserved motor learning in Alzheimer’s disease: Implications for anatomy and behavior. Journal of Neuroscience 6: 3006–9
- Fleischman D A, Gabrieli J D E 1998 Repetition priming in normal aging and Alzheimer’s disease: A review of findings and theories. Psychology and Aging 13: 88–119
- Gallagher M, Rapp P R 1997 The use of animal models to study the effects of aging on cognition. Annual Re iew of Psychology 48: 339–70
- Golomb J, de Leon M J, Kluger A, George A E, Tarshsh C, Ferris S H 1993 Hippocampal atrophy in normal aging: An association with recent memory impairment. Archives of Neurology 50: 967–73
- Jack C R Jr., Petersen R C, Xu Y C, Waring S C, O’Brien P C, Tangalos E G, Smith G E, Ivnik R J, Kokman E 1997 Medial temporal atrophy on MRI in normal aging and very mild Alzheimer’s disease. Neurology 49: 786–94
- Morrison J H, Hof P R 1997 Life and death of neurons in the aging brain. Science 278: 412–19
- Squire L R 1992 Memory and the hippocampus: A synthesis from findings with rats, monkeys, and humans. Psychological Re iew 99: 195–231
- West M J, Coleman P D, Flood D G, Troncoso J C 1994 Differences in the pattern of hippocampal neuronal loss in normal ageing and Alzheimer’s disease. The Lancet 344: 769–72
- Woodruff-Pak D S, Finkbiner R G 1995 Larger nondeclarative than declarative deficits in learning and memory in human aging. Psychology and Aging 10: 416–26
- Woodruff-Pak D S, Goldenberg G, Downey-Lamb M M, Boyko O B, Lemieux S K 2000 Cerebellar volume in humans related to magnitude of classical conditioning. NeuroReport 11: 609–15
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality