
Sample Neural Basis Of Learning And Memory Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
1. Basic Learningand Memory-Related Neural Changes
Much of our current understanding of basic memoryrelated neuronal changes and modifications stems from the pioneering research of Eric Kandel and his co-workers (Kandel 1998) who largely used an invertebrate, the marine mollusc Aplysia for their studies. Initially, they investigated simple memory mechanisms such as sensitization and habituation and found that there are both shortand long-term forms hereof and that the physiological correlates of the short-term forms are facilitated neuronal responses (increases in the magnitude of the excitatory postsynaptic potentials formed between sensory and motor neurons), and that of the long-term forms (repeated stimulus applications) required new ribonucleic acid (RNA) and protein synthesis, involving the growth of new synaptic connections between sensory and motor neurons. Later, they extended their work to gene expression and transcription factors which provided the genetic switch for memory consolidation and storage. A specific protein, named CREB, which stands for cAMP-responsive element binding protein, became a new standard label within this kind of research. cAMP stands for cyclic-adenosine monophosphate, a substance important for providing energy to cells. Other workers from both the invertebrate and vertebrate side demonstrated, for example, that adenylyl cyclase is an important enzyme for short-term memory (Zars et al. 2000) and long-term potentiation (LTP) as well as long-term depression (LTD). These are important mechanisms for memory consolidation (Lynch 2000). There is evidence that LTP and LTD lead to changes in the morphology of nerve cells, particularly to so-called synaptic plasticity (Coussens and Teyler 1996) and to changes in dendritic spine density (Engert and Bonhoeffer 1999).
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
The crucial questions are whether these findings can be directly transmitted to mammals and whether they hold for different kinds of memories (Eichenbaum et al. 1999). Here is less safe ground from current research (see, e.g., the findings of Zamanillo et al. [1999] against LTP as an essential prerequisite for memory formation) and the focus lies more strongly on metabolic changes which can be related to brain circuitries.
2. Memory Representation Within The Mammalian Nervous System
Amnesias in humans occur after a wide range of disease conditions (Markowitsch 2000). This fact alone implies that information processing in the mammalian brain is not controlled by a single center but, instead, by a distributed network. Furthermore, memory is also dependent on a large number of sensory, perceptual, attentive, emotional, and motivational processes each with its own anatomical substrates. The investigation of brain-damaged individuals consequently is of only limited predictive value for the action of the normal brain, as it demonstrates only what the brain can or cannot do without a particular region (Sometimes, a brain with a damaged region is compared with a chain in which a link is missing; this implies that with the lesioning approach the functions of the damaged area are, in a way, never attainable, as the respective region is missing). Therefore, the complementary approach of studying the intact brain with neuromonitoring (EEG, evoked potentials, magnetoencephalography) and the functional brain imaging methods methods ( positron emission tomography, functional magnetic resonance imaging, single photon emission tomography) provides the necessary completion for inferences on the action of the brain.
In spite of the present-day range of methodological approaches for studying learning and memory representations in the brain, the old dichotomy between localizationists and antilocalizationists remains and can be perceived as one of ideology. The localizationists have a more mosaic-like view of information representation, they speak of ‘centers’ and ‘modules,’ the antilocalizationists prefer a Gestalt-like view and emphasize interactions and networks. A bridge between the two can be built by assuming a mixture of hierarchical and parallel processes of information representation (McClelland and Rumelhart 1986; Barnden and Pollack 1991).
3. Memory Systems And Their Functional And Anatomical Correlates
The study of mammalian and in particular of human memory has to acknowledge the existence of several memory systems and their different neural representation. Aside from short-term memory, there are at least four major long-term memory systems: (a) the perceptual representation system (PRS), (b) procedural memory, (c) the knowledge system (or semantic memory), and (d) episodic memory (Schacter et al. 2000). The PRS refers to the fact that nonconscious presentation of a stimulus may lead to a higher probability of reidentification of it at a later time point; procedural memory is usually skill based, examples are riding a car, skiing, or playing the trumpet—that is, nonconscious, automatic processing of information. Both of these systems, but sometimes also the knowledge system, are termed implicit memory. The knowledge system (semantic memory) refers to general facts (‘world knowledge’) of the kind: ‘Madrid is the capital of Spain’ or ’12 is 144.’ While the knowledge system consists of context-free facts, the episodic memory system represents context (time, locus) embedded information. These episodes allow a mental time travel. One of the important distinctions between semantic and episodic memory is therefore that the latter allows to remember the episode and the former simply to know the fact.
3.1 Short-Term And Working Memory
The concept of working memory derived from the earlier one of short-term (and long-term) memories. While, however, short-term memory is seen as the online holding of information for an initial time period after first confrontation with it, working memory refers to a multicomponent, active processing of information which also includes the transmission of already long-term stored information in a temporary buffer prior to retrieval, and which is composed of a controlling central executive system and a number of subsidiary slave systems (Baddeley 2000).
Numerous experiments using monkeys, cats, and rats have reported that bilateral damage to the prefrontal cortex interferes with the short-term storage of information and that single units in this region display working memory related neuronal firing (Goldman-Rakic et al. 2000). Results from evokedpotential recordings and functional imaging studies have confirmed the dominant role of prefrontal regions for working memory in the human brain. Some reports found that damage to parietal regions impairs the on-line processing of information. Consequently, a frontoparietal network is seen as engaged in the control of attention and in processes of on-line working with memory.
3.2 The PRS And Procedural Memory
The two principal memory systems, which function independent of limbic system structures and of conscious reflections, are procedural memory and priming. There are two basic forms of priming, perceptual and conceptual—perceptual priming refers to the direct match between the presently and the previously perceived stimulus, while conceptual priming refers to the same category or class of the present and the previously perceived stimuli. The distinction between these two forms is important as conceptual priming apparently is more complexly represented in the brain, recruiting more or other neural structures than perceptual priming. Consequently, conceptual priming deteriorates in age-related brain diseases and possibly even in patients with limbic-system related amnesias, while perceptual priming does not. Conceptual priming has been related to prefrontal and even partly to medial temporal regions, while perceptual priming has much stronger affinities to unimodal areas of the posterior neocortex.
The procedural memory system is closely related to motor regions of the brain—to the basal ganglia, to portions of the frontal lobe, particularly of the premotor and motor cortex, and to cerebellar structures (some authors question, however, a specific role of cerebellar structures in procedural memory). Therefore, diseases such as Parkinson’s disease are taken as models—at least during the initial disease process—for selective procedural memory deficits (Thomas-Ollivier et al. 1999).
While not much is known about the processes from information encoding to information retrieval, there is at least preliminary evidence that principally the same structures activated during the initial stages of information transfer are also relevant for the late ones and even hold for storage of this implicitly learned information.
3.3 Brain Regions Relevant To Episodic Memory And The Knowledge System
Episodic memory and the knowledge system are sometimes combined and then named declarative memory (Squire and Knowlton 1999). Both systems have in common that they—at least during the stages of memory encoding and consolidation—rely on structures of the limbic system (Mesulam 2000), the episodic memory system most likely in addition on prefrontal structures. For retrieval of information, there seems to be a distinct lateralization; it appears that primarily right-hemispheric regions are engaged in episodic and primarily left-hemispheric ones in semantic-memory retrieval (Tulving et al. 1994, Markowitsch 2000).
The structures of the limbic system have long been discussed as important for transferring information from shortto long-term memory. More recently, components of this system have been combined under the headings ‘medial temporal lobe’ and ‘medial diencephalic memory system’ and have been named bottlenecks for information transmission from short to long-term memory (Markowitsch 2000). For the first system, the hippocampal formation was (and still is) seen as its hub for the second, medial thalamic nuclei and the hypothalamic mammillary bodies are core structures and Korsakoff’s syndrome is one of the most well-known diseases associated with it. However, it is more a matter of philosophical preferences whether one sees both systems as separate entities or as one. Likewise, there has been a waxing and waning discussion of the contributions of individual regions to memory formation (Tulving and Markowitsch 1998). Aside from the hippocampal region, the amygdala and paralimbic cortices have to be listed for the medial temporal system, and certain medial and anterior thalamic nuclei, the mammillary bodies, and interconnecting fiber bundles for the medial diencephalic one; structures of the basal forebrain and the ventral striatum may be added (Mesulam 2000, Markowitsch 2000).
There is another division of the limbic system which receives support from a number of sources, namely the distinction between two interacting circuits: (a) the medial (or Papez-) circuit centered around the hippocampus, and (b) the basolateral limbic circuit (amygdaloid circuit). While Papez (cited in Markowitsch 2000) considered his proposed circuit (hippocampal formation→ fornix→ mammillary bodies mammillothalamic tract → anterior thalamus → thalamocortical→peduncles → cingulate cortex→cingulum hippocampal formation) as principally engaged in the analysis of emotions, subsequent work showed that it plays a critical role for the transfer of information into long-term memory. The basolateral circuit is more closely related to emotional processing, but is also relevant for encoding the emotional valence of experiences. It includes the amygdala, the mediodorsal nucleus of the thalamus and associated regions, largely situated in the basal forebrain and in its extensions to anterior insular and temporopolar cortical areas plus interconnecting fibers such as the ventral amygdalofugal pathway, the anterior thalamic peduncle and the diagonal band (Fig. 1).
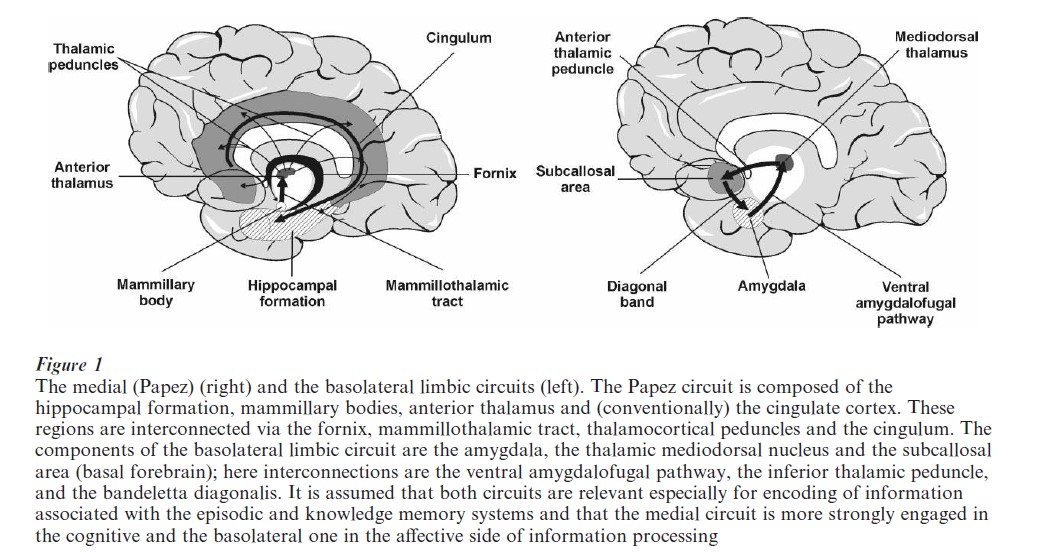
Bilateral damage to structures within the medial circuit is usually followed by severe memory disturbances—a fact which led researchers to speculate that amnesia might arise from an interruption of interconnections within this circuit and that the structures of this circuit are necessary for consolidation and for the transfer from shortto long-term storage. Bilateral damage to structures within the basolateral limbic circuit, on the other hand, seem to degrade memory processes by disconnecting the emotional from the mnestic properties of a given set of stimuli. For example, bilateral damage to the amygdaloid region may induce the Kluver–Bucy syndrome characterized by hyperorality, hyperphagia, agnosia, amnesia, hypersexuality, tameness placidity, and hypermetamorphosis. Since information encoding is, especially for episodic (autobiographic) information, dependent on emotional content, damage to the amygdala is likely to have detrimental effects on memory. This indeed was shown in patients with Urbach–Wiethe disease; such patients manifest a selective bilateral damage (mineralization) of the amygdalae. Two such patients were presented stories containing emotionally charged items and asked to recall them. They were significantly impaired because they could not show the preferential encoding of emotionally-laden vs. neutral items (Cahill et al. 1995).
Lesions in the basal forebrain (e.g., medial septum, basal nucleus of Meynert) may also disturb memory processing, though the deteriorations on the memory side are usually less severe and enduring than those after medial temporal or medial diencephalic damage; personality disturbances and tendencies to confabulate, on the other hand, are more common.
The similarity between the amnesic conditions arising after bilateral damage to medial temporal and medial diencephalic structures led Squire et al. (1993) to conclude that these structures might be components of a common memory system and to speak of ‘medial temporal–diencephalic amnesia.’ While this seems likely, there may also be regional functional specializations. Patients with bilateral diencephalic damage seem to have less awareness of their memory, a higher tendency to confabulate, and a larger span of retrograde amnesia than those with medial temporal lobe damage.
3.3.1 Encoding And Consolidation. While encoding refers to the more early stage of information acquisition, there are different views on the beginning and end of the consolidation phase. Sometimes, it is seen as directly succeeding the encoding phase, sometimes it is viewed as a process occurring after information has already been stored long term. As both encoding and consolidation of episodic and semantic memories engage structures of the limbic system, they can be perceived as closely related.
There are nevertheless single cases of patients with amnesias of various origins showing a wide range of unstable memory processing periods (Markowitsch 2000) so that it has to be concluded that there are several states and stages of memory consolidation which depend on complex anatomico–behavioral interdependencies. A comparison with repeated at- tempts of producing a complex painting may be apt. Still, medial temporal and diencephalic regions contribute substantially to both the stages of encoding and consolidating by triggering, accompanying, and assisting the steps from initial binding to the final assignment. Especially the hippocampus has been proposed to index ( previously encoded) information and assign it to specific cortical loci. Amygdaloid and septal regions may evaluate and filter for importance—that is, they screen the biological and social significance of perceived stimuli.
Aside from the more ‘classic’ limbic regions, functional brain imaging results have pointed to a strong role of prefrontal cortical areas in both encoding and retrieving of information. Among the first studies were those of Tulving and co-workers (1994) who condensed their findings in what they termed the ‘hemispheric-encoding-retrieval-asymmetry’ (HERA) model of information processing. This model considers prefrontal regions as important for information processing, but assigns different roles to the left and right frontal lobes: the left one being necessary for encoding of semantic and episodic information and the right one for the retrieval of episodic memories. While this model has sometimes been questioned or modified, there is still accumulating evidence for its principal validity. Current research tries to specify whether there are differential contributions of dorsolateral, polar, and inferolateral prefrontal regions and to what degree the prefrontal contribution can be regarded as a process of working memory.
3.3.2 Storage Of Information. The ways in which information is—frequently life-long—stored in the brain, is one of the biggest riddles of the neurosciences. Most scientists agree that the nervous system contains or forms the medium for storage, the more concrete mechanisms vary between species— invertebrates with their more ‘rudimentary’ nervous system may indeed have adopted other representations than mammals. Genetic and biochemical processes may nevertheless be common denominators (Kandel 1998). For mammalian nervous systems, information may very likely be distributed along widespread cerebral cortical networks (McClelland and Rumelhart 1986) with or without auxiliary engagement of subcortical limbic structures (Markowitsch 2000). The expanded cortical association areas with their large number of neurons and neuronal connections are primary candidates for information representation. Especially for emotionally colored memories—such as our biographical episodes—the cortical nets may engage and trigger affect-coding limbic regions for their rerepresentation or ‘ecphory’ (Markowitsch 2000, Sara 2000, Tulving and Markowitsch 1998). (Tulving reintroduced the term ecphory to describe the process by which retrieval cues interact with stored information so that an image or a representation of the information in question appears.)
There are, however, a few scientists who are of the opinion that most of our memories are represented outside our nervous system. Romijn (1997) considers a submanifest order of being as the principal storage place of memories. He is of the opinion that the whole universal in its spatial–temporal configuration would be permanently present in the submanifest order of being. Of this, a small portion would continuously materialize and thereby become available for psychic processing. Declarative memory would be found somewhere in the individual area of the submanifest order of being. Similar views are held by Sheldrake (1988) and Hameroff et al. (1998).
3.3.3 Retrieval Of Information. The conventional view is that retrieval activates similar (namely mainly limbic) brain structures as does encoding. This view may, however, be false or at least only partly correct: As mentioned above in connection with the HERA model of Tulving et al. (1994), the prefrontal cortex is a primary region for information retrieval. Converging evidence from brain-damaged patients and from studying normal subjects with functional imaging methods points to the necessity of (inferolateral) frontal and anterolateral temporal cortical regions for triggering the network-like stored information for successful retrieval (Markowitsch 1995, 2000) (Fig. 2). While a few authors even assume the temporal cortex to be the actual storehouse of information, here the view is favored that it provides the most direct access to the cortical network, while the prefrontal cortex is responsible for providing the effort necessary for a successful overall neural activation. Limbic structures of the medial diencephalon and the medial temporal lobe may have supporting roles in context-rich information retrieval (Markowitsch 2000).
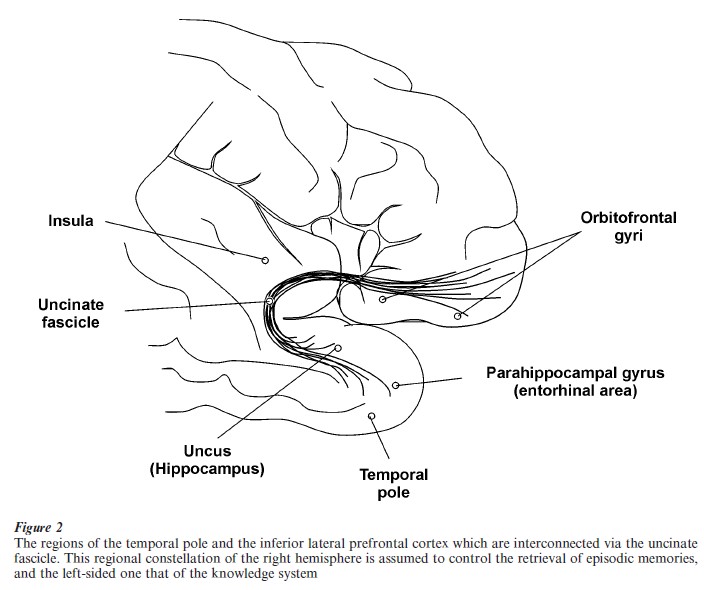
Retrograde amnesia—the failure of ecphory—may occur as a consequence of a progressively decaying cortical network—as is the case in patients with cortical dementia (e.g., Alzheimer’s disease) or, some- times, after a condition of severe hypoxia (e.g., after a heart attack and coma state). In this group of patients, the information is lost permanently. When the damage is restricted to the hippocampus and related structures in the medial temporal lobes, the retrograde amnesia may be temporally limited (covering a period of weeks in rodents, up to years or even decades in humans, leaving remote memories intact), suggesting to several authors that the hippocampus may play a role in the initial formation of new memories, and may somehow contribute to the gradual integration of new memories into neorcortical memory circuits (Squire et al. 1984, McClelland et al. 1995).
Alternatively, retrograde amnesia may be the consequence of a disrupted access to the net. Here, the expression ‘mnestic block syndrome’ has been coined to describe this phenomenon (Markowitsch 2000) which can be found both after manifest brain damage in the frontotemporal cortex and after so-called functional amnesia which is seen in patients with dissociative personality states. Such functional amnesias especially appear as a consequence of stress and trauma experiences. In this second category of patients, the mnestic block is usually selective, affecting primarily emotion-laden personal episodes, and it is assumed that the information— the engram—is in principle still manifest in the cortical nets.
Furthermore, static and functional imaging results from both neurologic and psychiatric patients point to alterations in especially the right temporofrontal regions when autobiographic information retrieval is disrupted and to corresponding changes in the left temporofrontal regions when semantic retrieval is impaired (Markowitsch 1995, 2000).
4. The Learning And Memory Networks—Conclusions
Learning and memory engage a wide range of brain regions—nevertheless, a conclusion that it is the total mass of the nervous system which codes information for storage would be a false one. Instead, the principles of convergence and divergence of information transmission hold throughout the nervous system and there are both modality and material-specific channels for information encoding, storage, and even retrieval and there are bits of information which are rich in affect and consciously reflected (and consequently require wide neural nets), while others are processed only subliminally and may never gain conscious representation. Furthermore, information is not represented in a static manner within the brain – mood, the environment, focused attention, internal associated cues, hormones, and many additional factors influence, modify, and rearrange our knowledge. The phenomena of state-dependent learning and retrieval and of false memories are examples for the susceptibility and vulnerability of our memories. The concept of plasticity has gained considerable support from invertebrate to human research and also the concept of life-long learning, proposed by the social sciences, demonstrates that information representation is much less hard-wired and rigid than formulated by traditional frameworks.
Bibliography:
- Baddeley A 2000 Short-term and working memory. In: Tulving E, Craik F I M (eds.) The Oxford Handbook of Memory. Oxford University Press, New York, pp. 77–92
- Barnden J A, Pollack J B (eds.) 1991 Advances in Connectionist and Neural Computation Theory: High-Level Connectionist Models. Ablex, Norwood, NJ, Vol. 1
- Cahill L, Babinsky R, Markowitsch H J, McGaugh J L 1995 The amygdala and emotional memory. Nature 377: 295–96
- Coussens C M, Teyler T J 1996 Long-term potentiation induces synaptic plasticity at nontetanized adjacent synapses. Learning and Memory 3: 106–14
- Eichenbaum H B, Cahill L F, Gluck M A, Hasselmo M E, Keil F C, Martin A J, McGaugh J L, Murre J, Myers C, Petrides M, Roozendaal B, Schacter D L, Simons D J, Smith W C, Williams C L 1999 Learning and memory: Systems analysis. In: Zigmond M J, Bloom F E, Landis S C, Roberts J L, Squire L R (eds.) Fundamental Neuroscience. Academic Press, San Diego, CA, pp. 1455–86
- Engert F, Bonhoeffer T 1999 Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature 399: 66–70
- Goldman-Rakic P S, Scalaidhe S E O, Chafee M V 2000 Domain specificity in cognitive systems. In: Gazzaniga M S (ed.) The New Cognitive Neurosciences. MIT Press, Cambridge, MA, pp. 733–42
- Hameroff S R, Kaszniak A W, Scott A C 1998 Toward a science of consciousness II: The second Tucson discussions and debates. The Tao of consciousness. Journal of Cognitive Neuroscience 11: 132–4
- Kandel E R 1998 A new intellectual framework for psychiatry. American Journal of Psychiatry 155: 457–69
- Lynch G 2000 Memory consolidation and long-term potentiation. In: Gazzaniga M S (ed.) The New Cognitive Neurosciences, 2nd edn. MIT Press, Cambridge, MA, pp. 139–58
- Markowitsch H J 1995 Which brain regions are critically involved in the retrieval of old episodic memory? Brain Research Reviews 21: 117–27
- Markowitsch H J 2000 Memory and amnesia. In: Mesulam MM (ed.) Principles of Cognitive and Behavioral Neurology. Oxford University Press, New York, pp. 257–93
- McClelland J L, McNaughton B L, O’Reilly R C 1995 Why there are complementary learning systems in the hippocampus and neocortex: Insights from the successes and failures of connectionist models of learning and memory. Psychological Review 102: 419–57
- McClelland J L, Rumelhart D E 1986 A distributed model of human learning and memory. In: McClelland J L, Rumelhart D E, and the PDP Research Group (eds.) Parallel Distributed Processing. Explorations in the Microstructure of Cognition: Psychological and Biological Models. MIT Press, Cambridge, MA, Vol. 2, pp. 171–215
- Mesulam M-M 2000 Behavioral neuroanatomy. Large-scale networks, association cortex, frontal syndromes, the limbic system, and hemispheric specializations. In: Mesulam M-M (ed.) Principles of Cognitive and Behavioral Neurology. Oxford University Press, New York, pp. 1–120
- Page M 2000 Connectionist modelling in psychology: A localist manifesto. Behavioral and Brain Sciences 23: 443–512
- Romijn H 1997 About the origin of consciousness. A new, multidisciplinary perspective on the relationship between brain and mind. Proceedings an de Koninklijke Nederlandse Akademie an de Wetenschappen 100: 181–267
- Sara S J 2000 Retrieval and reconsolidation: Toward a neurobiology of remembering. Learning and Memory 7: 73–84
- Schacter D L, Wagner A D, Buckner R L 2000 Memory systems of 1999. In: Tulving E, Craik F I M (eds.) The Oxford Handbook of Memory. Oxford University Press, New York, pp. 627–44
- Sheldrake R 1988 The Presence of the Past: Morphic Resonance and the Habits of Nature, 1st edn. Times Books, New York
- Squire L R, Cohen N J, Nadel L 1984 The medial temporal region and memory consolidation: A new hypothesis. In: Weingartner H, Parker E (eds.) Memory Consolidation. Erlbaum, Hillsdale, NJ, pp. 185–210
- Squire L R, Knowlton B J 1999 The medial temporal lobe, the hippocampus, and the memory system of the brain. In: Gazzaniga M S (ed.) The New Cognitive Neurosciences, 2nd MIT Press, Cambridge, MA, pp. 765–79
- Squire L R, Knowlton B, Musen G 1993 The structure and organization of memory. Annual Reviews of Psychology 44: 453–95
- Thomas-Ollivier V, Reymann J M, Le Moal S, Schuck S, Lieury A 1999 Procedural memory in recent-onset Parkinson’s disease. Dementia and Geriatric Cognitive Disorders 10: 172–80
- Tulving E, Kapur S, Craik F I M, Moscovitch M, Houle S 1994 Hemispheric encoding retrieval asymmetry in episodic memory: Positron emission tomography findings. Proceedings of the National Academy of Sciences of the USA 91: 2016–20
- Tulving E, Markowitsch H J 1998 Episodic and declarative memory: Role of the hippocampus. Hippocampus 8: 198–204
- Zamanillo D, Sprengel R, Hvalby O, Jensen V, Burnashev N, Rozov A, Kaiser K M, Koster H J, Borchardt T, Worley P, Lubke J, Frotscher M, Kelly P H, Sommer B, Andersen P, Seeburg P H, Sakmann B 1999 Importance of AMPA receptors for hippocampal synaptic plasticity but not for spatial learning. Science 284: 1805–11
- Zars T, Fischer M, Schulz R, Heisenberg M 2000 Localization of a short-term memory in Drosophila. Science 288: 672–5
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality