
Sample Neural Basis Of Neglect Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
1. The Term ‘Hemispatial Neglect’
One of the most dramatic deficits in spatial perception and action is hemispatial neglect (neglect for short), a neurobehavioral disorder in which a patient is unable to acknowledge, report, or otherwise make explicit use of information falling in the visual hemispace contralateral to the responsible lesion (Bartolomeo and Chokron 2001, Bisiach and Vallar 2001, Driver 1998, Vallar 1998). The most frequently affected neural region associated with neglect is the inferior parietal lobule although other cortical and subcortical sites have also been implicated (Karnath et al. 2001). Neglect occurs more commonly and with greater severity after right than left-hemisphere lesions in humans and the differential involvement of the right hemisphere in processes of attention and spatial representation is consistent with recent neuroimaging studies (Corbetta et al. 2000). Because of this asymmetry, neglect will be referred to as ‘left-sided’ throughout this section.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1.1 The Phenomenon Of Hemispatial Neglect
Neglect can be severe and debilitating in everyday life; individuals may fail to notice objects on the left of a scene, may ignore words on the left of a page or food on the left of a plate, and typically, may omit to copy features on the left of a figure while preserving the corresponding features on the right, as shown in Fig. 1 Neglect is also standardly elicited in tasks that require the patient to cross out items that appear randomly scattered on a piece of paper or to mark the midpoint of a single horizontal line.
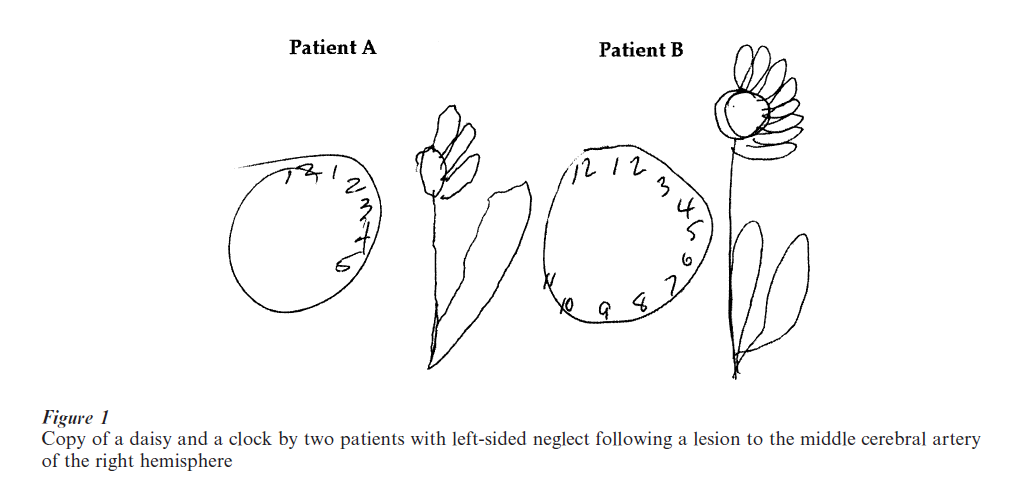
Neglect is not restricted to the visual domain; patients may not orient to contralateral auditory information, may not detect a contralateral somatosensory stimulus and may even ignore contralateral odors. Neglect is also not restricted to sensory input; patients with neglect may plan and even execute movements more poorly to the contralateral than ipsilateral side and this holds both for upper limb movements (Heilman et al. 1987, Mattingley and Driver 1997, Mattingley et al. 1998) as well as for eye movements (Gainotti 1993). The relative paucity of eye movements to the left is illustrated in Fig. 2 in which we see, relative to control subjects, both fewer and more brief eye fixations contralesionally as well as a simultaneous increase in the number and duration of right-sided fixations.
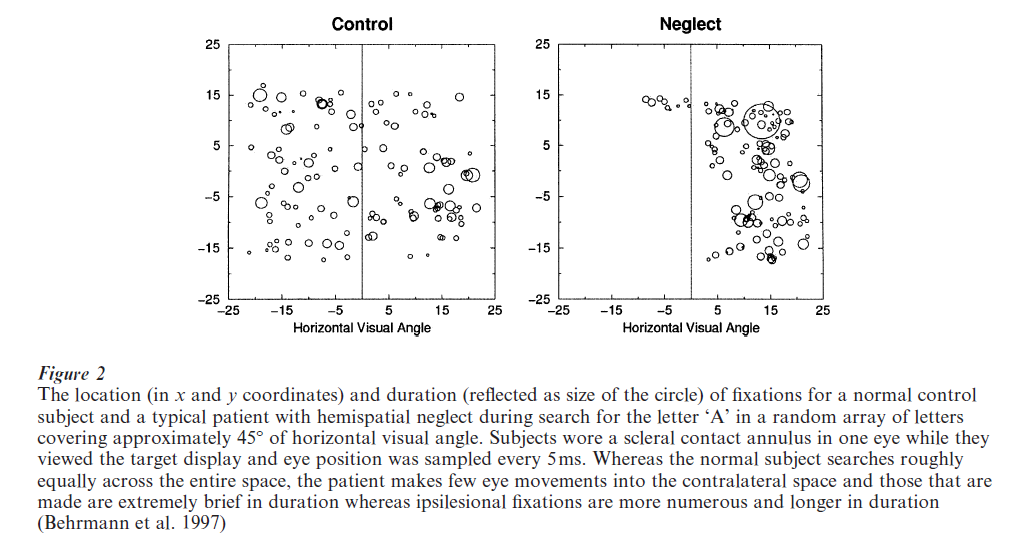
The failure to process information on the contralateral side cannot be attributed to a primary motor impairment nor to a hemianopia; even when information is presented to the intact visual field of a neglect patient, information in the contralateral hemispace is still neglected (Ladavas et al. 1990). Patients may even show neglect in the absence of visual input; for example, patients ignore the left side of a scene constructed in visual mental imagery (Bisiach and Luzzatti 1978) and fail to search the left of a dark room (Hornak 1992). Finally, patients can be cued to process information on the contralesional side (Milner et al. 1993, Riddoch and Humphreys 1983) through verbal or other visual instructions. Taken together, these findings all rule out a primary sensory deficit as being causally linked to neglect although a field defect can exacerbate any tendency towards neglect (Doricchi and Angelelli 1999).
Neglect occurs most frequently along the horizontal plane although ‘altitudinal neglect,’ in which information appearing in the upper rather than lower portion of an array (or vice versa) is ignored, may also occur. Neglect may also occur along the radial dimension although this is thought to be tied to the need for a motor response and is less commonly seen when perception alone is tested.
1.2 Related Disorders
There are a number of deficits related to neglect. ‘Extinction to double simultaneous stimulation’ refers to a phenomenon in which patients fail to report the stimulus on the contralesional side when two stimuli are presented simultaneously to the ipsilateral and contralateral side of space. When only a single stimulus is presented, even to the contralateral side, it can be detected and reported. Although some consider extinction to be a mild form of neglect, their exact relationship remains controversial. A number of recent studies have suggested that extinction arises from a competition for attention and activation between two stimuli (di Pellegrino et al. 1997); unilateral brain damage represents a reduction in the competitive weights on the affected side and, as a consequence, stimuli appearing on that side are only weakly activated and hence dominated by stronger ipsilateral stimuli. Extinction can be observed both when the two stimuli are from the same sensory modality (e.g., visual) or even when they are from two different modalities (e.g., visual and tactile) (also see Ladavas et al. (1998), and for related work in monkeys, Graziano and Gross (1998)).
A second, related phenomenon is ‘allesthesia,’ in which a patient is able to detect the contralesional stimulus but refers its position to the intact side. Asked where the stimulus had appeared, the patient usually locates the detected stimulus on the ipsilesional side. Finally, ‘anosagnosia,’ a disorder in which patients are unaware of their limited representation of the environment and consequently minimize or deny that they have any impairment, may co-occur with neglect and may adversely affect attempts at rehabilitation.
2. Underlying Mechanisms
A number of general interpretations have been advanced to account for the varied manifestations of neglect. For example, some have suggested that neglect arises from a deficit in orienting spatially to the contralesional side with a gradient of attention biasing attention to the ipsilateral side. Others have argued that the problem is one in which there is a rotation or translation of the egocentric frame of reference towards the ipsilesional side. Yet a third suggestion, the premotor theory of neglect, is that there is an imbalance between the spatial representations that control motor programs, and this imbalance gives rise to an attentional deficit. Although described as alternatives, these differing interpretations may not be mutually exclusive and may refer to different aspects of a single deficit. What is clear at present is that, although they may diverge in detail, these explanations share a common perspective and that is that neglect is a higher-order spatial impairment.
One view of neglect, formalized most recently by Pouget and Driver 2000) is that neglect reflects a partial loss or dysfunction of cells in the parietal cortex and other related neural areas. Because there is a certain number of neurons for representing information in different lateral positions, damage to these neurons gives rise to a pathological gradient with a smooth decline for increasingly leftward locations and an off-center peak reflecting the peak activation on the ipsilateral side. A gradient has been incorporated into several neural network models that simulate neglect and these models have successfully characterized many aspects of neglect including extinction, neglect on linebisection and line cancellation tasks, and neglect in reading (Mozer and Behrmann 1990, Mozer et al. 1997, Pouget and Sejnowski 1999).
3. Spatial Reference Frames
One question that has been addressed in the context of neglect concerns the nature of the spatial representations which, when damaged, give rise to the disorder. Specifically, the approach adopted to address this issue has involved determining what constitutes ‘left’ and ‘right.’ Thus, when the patients ignore information on the left, what is it left of? Because ‘left’ and ‘right’ cannot be defined in absolute terms, they must always be qualified with respect to a set of coordinates or spatial reference frame, i.e., relative to an origin and axes. It is these spatial coordinates that are used for organizing spatial information, and for establishing spatial relationships between objects in the scene and between the observer and the objects. By examining what reference frame(s) determines the midline such that information to the left of it is neglected, we may obtain an understanding of how spatial position is coded in the parietal cortex and associated regions.
A major distinction between the types of spatial reference frames has been between those that define spatial position egocentrically or with respect to the viewer and those that define position allocentrically, irrespective of the position of the viewer. At least four different egocentric frames have been identified, with the midline centered on: (a) the eyes or vertical meridian of the visual field, (b) the head, (c) the trunk, or (d) the longitudinal axis of the limb that is involved in executing an action, such as the arm. Other studies have shown that patients neglect information on the left, defined allocentrically, with respect to the gravitational upright or environment (or scene) or with respect to the midline of individual objects or perceptual units in the scene.
Several important insights concerning the spatial coordinates used for coding left and right have been obtained recently. Among these is the finding that spatial position may be coded simultaneously with respect to multiple frames of reference, both egocentric and allocentric. As depicted in Fig. 3, a patient, seated in front of a large screen, reports the identity of letters from a display. In the first condition (panel A), the patient’s head, eyes, and trunk are all aligned with the midline of the scene and the patient reports fewer letters from the left than right. Because, under this condition, the midlines of the different reference frames are all aligned, one cannot distinguish the relative contribution of these various egocentric and allocentric reference frames to the observed neglect. To deconfound this, in a second condition, the viewer’s midline is rotated out of alignment from the midline of the scene and now the upper and lower portions of the display coincide with the viewer’s left and right, respectively. Interestingly, in this second condition (panel B), the patient continues to report less information from the left than right of the midline of the allocentric scene-based frame but also reports fewer letters from the top than bottom of the display, implicating a viewer-based reference frame. Indeed, in that quadrant (upper left) in which both allocentric and egocentric neglect co-occur, there is the greatest neglect with the relationship between these two reference frames apparently being additive.
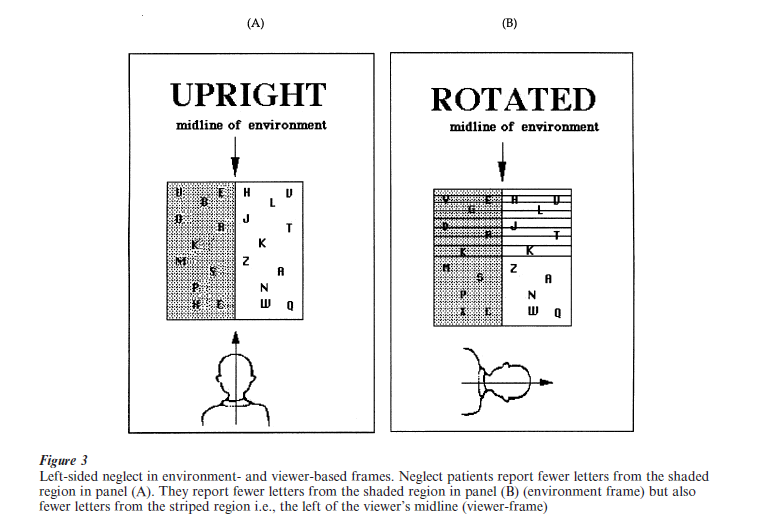
Several recent studies have also documented the presence of neglect defined with respect to the midline of individual objects in the scene. In one such study, neglect patients saw computer displays consisting of an object (barbell) in which the two circles were differently colored (Fig. 4, panel (A)) (Behrmann and Tipper 1994). The barbell appeared in the center of the computer screen (or entirely on the right for patients with a left field defect) and then, after a short delay, on two-thirds of the trials, a target or probe in the form of a small white dot appeared in the center of either the left or the right circle. The patients pressed a key as soon as they detected the presence of this target. As expected, in this ‘static’ condition, patients were poorer at detecting the left than the right target. Because the left and right of the barbell object are aligned with reference frames defined by the viewer and scene, a decoupling condition was introduced. In this second ‘rotating’ or moving condition, the midline of the barbell was dissociated from various other reference frames in the following way: first, the barbell was presented on the computer screen and then, after a short delay, it was rotated 180o around the midpoint of the connecting horizontal rod while the patient watched. When the rotation was complete and the barbell had reached its stationary position, a short delay ensued and then the target appeared on the left or right. Note that, as shown in Fig. 4, the display in which the target finally appears is identical in the static and rotating condition. However, in the rotation condition, the original left circle of the barbell is now on the right side of the computer screen and the original right circle is now of the left side of the screen. This arrangement allows us to determine whether the patients now neglect targets appearing on the ‘good’ right side of space (but on the left of the object) and show good detection of targets appearing on the ‘poor’ left side of space (but on the right of the object).
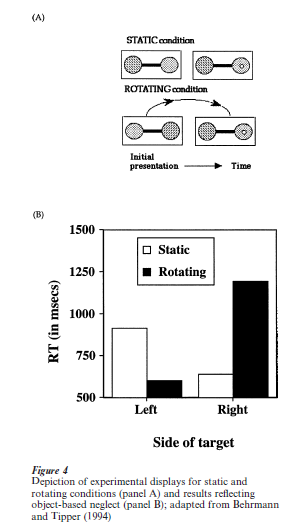
In contrast with the static condition, in the rotating condition, patients are faster at detecting the leftthan right-sided target (panel B). The explanation for this paradoxical reversal of neglect is that patients neglect the information that falls to the left of the object’s midline and, conversely, attend well to the information that falls to the object’s right. Even when the object moves into a new spatial location or undergoes a complete rotation, information on its left is still at a disadvantage compared with information on its right. This pattern cannot be attributed to an artifact of eye movement; even when eye movement is controlled, the same pattern is observed (Tipper and Behrmann 1996). It also is not obviously attributable to the covert tracking of the left or right of the object. In fact, relative to the static condition, the facilitation for targets on the left and inhibition of targets on the right in the rotation condition critically depend on the two circles forming a unified object and are not evident when the two circles are not joined. If the result were simply a function of the patients’ covertly tracking say the right circle of the barbell (to which their attention is drawn initially), a similar finding should be obtained even when the two circles are not connected. These findings lead to the conclusion that spatial positions are also defined with respect to an object-based reference frame and information to the left of this frame may be neglected. These findings are compatible with results from a single neuron recording study in monkeys which shows directional selectivity for neurons in the parietal cortex in a task in which saccades are made to the relative left or right of a stimulus bar (Olson and Gettner 1998).
Interestingly, this object-based neglect can occur simultaneously with neglect defined with respect to the environment or to the viewer (Behrmann and Tipper 1999). In an experiment using similar displays but with two grey squares appearing simultaneously with the barbell, one on the left and one on the right, and which remain stationary throughout all conditions, object based neglect is observed (as above, see Fig. 4). More importantly, detection of the probe in the left square is poorer than in the right square, revealing the presence of neglect for information on the left not only in an object-defined frame but also (and concurrently) in a frame defined by the midline of the viewer or scene. Finally, the extent of neglect in one of these reference frames can be exacerbated by presenting the probe more frequently within this one frame and less often in other reference frames. Taken together, these results suggest two important conclusions: first, patients can neglect the left of the environment and/or viewer and, at the very same time, can neglect the left of an object, even when the left of the object is on the right side of space (as demonstrated by the rotating barbell). Second, the distribution of visuospatial attention in these two frames of reference is flexible, and may be influenced by task demands or contingencies. These findings suggest that the parietal lobe can code spatial information relative to multiple spatial reference frames simultaneously and flexibly.
Studies concerned with reference frames have also been conducted using single unit recording techniques with awake, behaving monkeys either with or without lesions. Although there does not appear to be a direct analog of hemispatial neglect in monkeys, and anatomical and cytoarchitectonic homologies between the posterior parietal cortex in monkeys and humans are not perfect (Crowne and Mah 1998), several experimenters have described the occurrence of visual extinction after lesions to the posterior parietal areas (Gaffan and Hornak 1997, Lynch and McLaren 1989, Rizzolatti et al. 1985). Converging evidence is also obtained from recent neurophysiological studies with nonlesioned monkeys in which the spatial coordinates mediated by the parietal cortex are explored (for recent reviews see Andersen et al. 1997, Colby and Goldberg 1999).
4. Implicit Visual Information Processing
One of the most provocative claims in the research on neglect is that, despite the loss of phenomenal awareness of the contralateral information, these patients appear to have processed a considerable amount of contralateral information implicitly and this information can be used to guide their behavior (Driver and Vuilleumier 2001). An early observation by Kinsbourne and Warrington (1962) suggested that these patients ‘knew’ more than their overt responses suggested. Thus, in their reading errors, patients with neglect dyslexia often retained word length, for example, producing MILKMAN for MAILMAN. Since that time, several innovative paradigms have been developed to explore this issue. It has been shown, for example, that patients can make same different judgments about two pictures, one presented to each visual field, significantly above chance even though they are unable to identify the left-sided pictures, can make meaningful and consistent preference ratings on aspects on the left of stimuli even though they fail to report the presence of the information on which their apparent preference ratings were made, and can show facilitation in a priming task when the prime appears on the left and the probe centrally (McGlincheyBerroth 1997).
5. Perspectives On Rehabilitation
A diverse set of strategies have been adopted in studies which attempt to rehabilitate patients with neglect. These include training visual information and visual search, tactile exploration, actively or passively moving the contralesional arm and imagining mental scenes. Caloric vestibular stimulation in which iced water is introduced to the left ear and stimulates the vestibular apparatus has been shown to lead to a dramatic, albeit, short-term improvement in the disorder. Vibration of the left neck muscles (which produces an afferent that is similar to twisting the trunk midline leftward) has also been shown to be successful. Finally, a recent study, in which patients wore optical prisms that shifted the visual scene to the right, showed that patients were increasingly able to reach towards the contralesional side. The neural basis of the improvements remain poorly understood and may involve plasticity in some brain areas although the parietal lobe is notoriously nonplastic (Kolb and Cioe 1998). It is also possible, however, that whatever change is observed may arise from involvement of other postural alterations (such as the shift of the viewer-midline more leftward).
6. Concluding Remarks And Future Directions
As is clear from this review, hemispatial neglect is a heterogeneous and complicated disorder that defies simple explanation. For example, there remains a pressing need to understand what gives rise to this disorder, and what frames of reference are used to code spatial position. Additionally, we need to know how structured and unified behavior emerges if position is defined with respect to multiple coordinates. One possible explanation is that the contribution of each set of coordinates can be differentially weighted according to the task at hand. Consistent with this task dependency is the fact that the activity in neurons in the posterior parietal cortex differs depending on the task the animal is intending to perform (eye movements vs. reaching; Snyder et al. 1997). An additional pressing issue concerns the fate of the neglected information; whether this information is processed normally and in a rich and detailed fashion but is somehow not available for conscious or explicit report remains debatable and highly controversial. Promising studies using functional magnetic resonance imaging have shown differences in brain activation in the striate and extrastriate regions of the contralateral hemisphere for stimuli that have been extinguished compared with those that are reported (Driver and Vuilleumier 2001, Marzi et al. 2000, Rees et al. 2000, Vuilleumier et al. 2001).
A final issue to resolve concerns the role of the parietal temporal cortex in relation to other cortical regions that are involved in spatial processing. The parietal temporal cortex does not have a monopoly on spatial processing and regions, such as the frontal cortex and parahippocampal regions, also contribute importantly to spatial representations. How these different areas work together to give rise to our unitary visual experience remains unclear but neuropsychologic studies with patients suffering from hemispatial neglect allow us to observe some of the representations and processes that ultimately contribute to our unified spatial experience. These findings together with those obtained from functional imaging studies with normal subjects, single-unit recordings from nonhuman primates and computational analyses continue to provide new insights into this perplexing disorder.
Bibliography:
- Andersen R A, Snyder L H, Bradley D C, Xing J 1997 Multimodal representation of space in the posterior parietal cortex and its use in planning movements. Annual Review of Neuroscience 20: 303–30
- Bartolomeo P, Chokron S 2001 Levels of impairment in unilateral neglect. In: Boller F, Grafman J (eds.) Handbook of Neuropsychology. Elsevier Science, Amsterdam, Vol. 4, pp. 67–98
- Behrmann M, Barton J J S, Watt S, Black S E 1997 Impaired visual search in patients with unilateral neglect: An oculographic analysis. Neuropsychologia 35: 1445–58
- Behrmann M, Tipper S P 1994 Object-based attentional mechanisms: Evidence from patients with unilateral neglect. In: Umilta C, Moscovitch M (eds.) Attention and Performance XV: Conscious and Nonconscious Information Processing. MIT Press, Cambridge, MA, pp. 351–75
- Behrmann M, Tipper S P 1999 Attention accesses multiple reference frames: Evidence from neglect. Journal of Experimental Psychology: Human Perception and Performance 25: 83–101
- Bisiach E, Luzzatti C 1978 Unilateral neglect of representational space. Cortex 14: 129–33
- Bisiach E, Vallar G 2001 Unilateral neglect in humans. In: Boller F, Grafman J (eds.) Handbook of Neuropsychology. Elsevier Science, Amsterdam, pp. 459–502
- Colby C L, Goldberg M E 1999 Space and attention in parietal cortex. Annual Review of Neuroscience 22: 319–49
- Corbetta M, Kincade J M, Ollinger J M, McAvoy M P, Shulman G L 2000 Voluntary orienting is dissociated from target detection in human posterior parietal cortex. Nature Neuroscience 3: 1–6
- Crowne D P, Mah L W 1998 A comparison of hemispatial neglect from posterior parietal and periarcuate lesions in the monkey. Psychobiology 26: 103–8
- di Pellegrino G, Basso G, Frassinetti F 1997 Spatial extinction on double asynchronous stimulation. Neuropsychologia 35: 1215–23
- Doricchi F, Angelelli P 1999 Misrepresentation of horizontal space in left unilateral neglect: Role of hemianopia. Neurology 52: 1845–52
- Driver J 1998 The neuropsychology of spatial attention. In: Pashler H (ed.) Attention. Psychology Press, Hove, UK, pp. 297–340
- Driver J, Vuilleumier P 2001 Perceptual awareness and its loss in unilateral extinction. Cognition 79: 39–88
- Gaffan D, Hornak J 1997 Visual neglect in the monkey. Brain 120: 1647–57
- Gainotti G 1993 The role of spontaneous eye movements in orienting attention and in unilateral neglect. In: Robertson I, Marshall J C (eds.) Hemispatial neglect. Lawrence Erlbaum, London, pp. 107–22
- Graziano M S, Gross C G 1998 Spatial maps for the control of movement. Current Opinion in Neurobiology 8: 195–201
- Heilman K M, Bowers D, Valenstein E, Watson R T 1987
- Hemispace and hemispatial neglect. In: Jeannerod M (ed.) Neurophysiological and Neuropsychological Aspects of Spatial Neglect. Elsevier Science, Amsterdam, pp. 115–50
- Hornak J 1992 Ocular exploration in the dark by patients with visual neglect. Neuropsychologia 30: 547–52
- Karnath H O, Ferber S, Himmelbach M 2001 Spatial awareness is a function of the temporal not the posterior parietal lobe. Nature 411: 950–3
- Kinsbourne M, Warrington E K 1962 A variety of reading disability associated with right hemisphere lesions. Journal of Neurology, Neurosurgery and Psychiatry 25: 339–44
- Kolb B, Cioe J 1998 Absence of recovery or dendritic organization after neonatal posterior parietal lesions. Psychobiology 26: 134–42
- Ladavas E, di Pellegrino G, Farne A, Zeloni G 1998 Neuropsychological evidence of an integrated visuotactile representation of peripersonal space in humans. Journal of Cognitive Neurosciences 10: 581–9
- Ladavas E, Petronio A, Umilta C 1990 The deployment of visual attention in the intact field of hemineglect patients. Cortex 26: 307–17
- Lynch J C, McLaren J W 1989 Deficits of visual attention and saccadic eye movements after lesions of parietooccipital cortex in monkeys. Journal of Neurophysiology 61: 74
- Marzi M, Girelli M, Mimussi C, Smania N, Maravita A 2000 Electrophysiological correlates of conscious vision: Evidence from unilateral extinction. Journal of Cognitive Neuroscience 12: 869–77
- Mattingley J B, Driver J 1997 Distinguishing sensory and motor deficits after parietal damage: An evaluation of response selection biases in unilateral neglect. In: Thier P, Karnath H O (eds.) Parietal Contributions to Orientation in 3D Space. Springer, Heidelberg, Germany, pp. 309–38
- Mattingley J B, Husain M, Rorden C, Kennard C, Driver J 1998 Motor role of human inferior parietal lobe revealed in unilateral neglect patients. Nature 392: 179–82
- McGlinchey-Berroth R 1997 Visual information processing in hemispatial neglect. Trends in Cognitive Sciences 1: 91–7
- Milner A D, Harvey M, Roberts R C, Forster S V 1993 Line bisection errors in visual neglect: Misguided action or size distortion? Neuropsychologia 31: 39–49
- Mozer M C, Behrmann M 1990 On the interaction of selective attention and lexical knowledge: A connectionist account of neglect dyslexia. Journal of Cognitive Neuroscience 2: 96–123
- Mozer M C, Halligan P W, Marshall J C 1997 The end of the line for a brain-damaged model of hemispatial neglect. Journal of Cognitive Neuroscience 9: 171–90
- Olson C R, Gettner S N 1998 Impairment of object-centered vision following lesions of macaque posterior parietal cortex. Society of Neuroscience Abstracts 24: 1141
- Pouget A, Driver J 2000 Relating unilateral neglect to the neural coding of space. Current Opinion in Neurobiology 10: 242–9
- Pouget A, Sejnowski T 1999 A new view of hemineglect based on the response properties of parietal neurones. In: Burgess N, Jeffery K J, O’Keefe J (eds.) Spatial Functions of the Hippocampal Formation and Parietal Cortex. Oxford University Press, Oxford, UK, pp. 127–46
- Rees G, Wojciulik E, Clarkje K, Husain M, Frith C, Driver J 2000 Unconscious activation of visual cortex in the damaged right hemisphere of a parietal patient with extinction. Brain 123: 1624–33
- Riddoch M J, Humphreys G W 1983 The effect of cueing on unilateral neglect. Neuropsychologia 21: 589–99
- Rizzolatti G, Gentilucci M, Matelli M 1985 Selective spatial attention: One center, one circuit, or many circuits. In: Posner M I, Marin O S M (eds.) Attention and Performance XI. Lawrence Erlbaum Associates, Hillsdale, NJ, pp. 251–65
- Snyder L H, Batista A P, Andersen R A 1997 Coding of intention in the posterior parietal cortex. Nature 386: 167–70
- Tipper S P, Behrmann M 1996 Object-centred not scene-based visual neglect. Journal of Experimental Psychology: Human Perception and Performance 22: 1261–78
- Vallar G 1998 Spatial hemineglect in humans. Trends in Cognitive Sciences 2: 87–96
- Vuilleumier P, Sagiv N, Hazeltine E, Poldrack R A, Swick D, Ragal R D, Gabrieli J D E 2001 Neural fate of seen and unseen faces in visuo spatial neglect: A combined event-related functional MRI and event-related potential study. Proceedings of the National Academy of Sciences 98: 3495–3500
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality