
Sample Fear Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
Fear is an innate behavioral–cognitive–emotional system that evolved because of its evolutionary success in protecting animals (including humans) from danger. While fear to certain kinds of stimuli is unlearned, fear can also be acquired rapidly and lastingly to other kinds of stimuli, allowing animals to respond adaptively to new environmental threats. This research paper will present some of the behavioral and physiological aspects of fear that make it a useful model system for understanding the psychological and neurobiological mechanisms of emotion, learning, and memory, as well as the development of a variety of human fear-related disorders.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Fear—Definition And Background
All organisms are subject to threats and dangers from their environments, such as predation, dominance, and territoriality. Through the process of natural selection, evolution has endowed all organisms with defensive mechanisms (e.g., chemical toxins in plants; fight-flight responses in animals) for self-preservation. Fear serves this same purpose. In humans and other higher functioning animals, ‘fear’ as a scientific term describes a behavioral–cognitive–emotional state in which a set of adaptive responses is activated in the presence of danger. As the term is more commonly used, ‘fear’ refers to a sensation (or feeling) of strong unpleasant emotion aroused by stimuli or events that are perceived as threatening or dangerous.
While this more anthropomorphic ( psychological) sense of the word ‘fear’ may be useful in describing a person who is experiencing an environmental threat (e.g., electric shock) and exhibiting certain stereotyped behaviors (such as a fearful facial expression, autonomic arousal, behavioral readiness, and other defensive responses), it is probably inaccurate to use the term to describe a goldfish (or any other lower functioning animal) that is exhibiting similarly stereotyped behaviors in response to electric shock (such as avoidance or escape). In this research paper, fear will be defined as a by-product (or an emergent property) of a coordinated pattern of brain activities and bodily changes that appear in response to threats and dangers. Thus, fear serves an adaptive function, but evolved relatively recently among the continuum (or phylogeny) of defensive mechanisms.
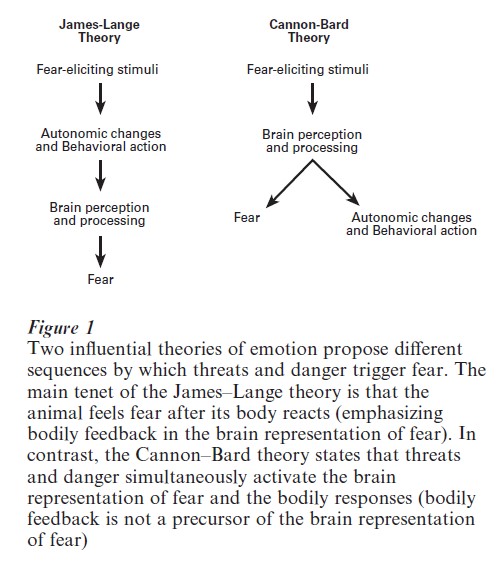
Fear as an emotion has been a topic of interest to many subdisciplines of psychology, including psychoanalysis, behaviorism, cognitive psychology, and biological psychology (for detailed treatment and further references, see LeDoux 1996). Historically, two opposing theories of emotion, the James–Lange theory and the Cannon–Bard theory, have been especially influential (Fig. 1). According to the James– Lange theory, threatening or dangerous stimuli trigger physiological arousal and behavioral responses (via the autonomic nervous system), which are perceived and then processed as fearful by the brain (emphasizing the importance of bodily feedback to the brain). In contrast, the Cannon–Bard theory posits that threats and dangers simultaneously trigger brain activities representing fear and initiate physiological arousal and behavioral readiness (emphasizing the importance of the brain activity). Thus, the critical distinction between the two theories is whether the appearance of immediate danger (e.g., a bear) is postulated to activate the central fear system in parallel with fleeing and autonomic responses (Cannon–Bard theory), or to first trigger fleeing and autonomic responses, which are then detected and interpreted as fear by the brain (James–Lange theory). There is evidence in support of both theories, suggesting that central processes and bodily feedback (as well as other, cognitive processes) both contribute to the emergence of fear as an emotion.
2. Neuronal Substrates Of Fear
Because humans share many biological and behavioral features with other animals, experimental studies with nonhuman animals provide valuable information toward understanding the physiological as well as the psychological aspects of fear. Darwin postulated in The Expression of Emotions in Man and Animals (1872) that emotions such as fear are innate, highly specific to situations, and the product of coordinated brain activities, and that different animal species with diverse environmental demands (or histories) have evolved dissimilar specialized sets of fear (or defensive) responses to maximize survival. It follows from this line of reasoning that more related species will exhibit more similar fear responses than will less related species, whose nervous systems share fewer characteristics. Thus, it is highly likely that the neuronal substrates underlying fear (or other defensive mechanisms) in humans are more similar to those of other mammals (e.g., chimpanzees and rats) than those of nonmammalian animals (e.g., fish and birds).
A repertoire of innate fear responses that is characteristic of a particular animal species is often referred to as species-specific defensive reactions (SSDRs) (Bolles 1970) (Fig. 2, A). For instance, the universal expressions of fear in humans include particular facial expressions (seen in members of all cultures), changes in autonomic nervous system activities (e.g., increased heart rate, increased blood pressure, rapid respiration, perspiration, elevated pain threshold, pupil enlargement, and decreased digestive activity) and behavioral readiness (e.g., freezing, increased propensity to startle, and sharpening of cognition) (see LeDoux 1996). These fear responses in humans are ‘hardwired’ (innate) as indicated by the observation that newborn infants, when exposed to aversive stimuli (e.g., a loud noise) for the very first time, exhibit responses, such as sudden eye closure, rapid breathing, clutching of hands, and crying, which are indicative of a general state of fear (e.g., Watson 1924). The innateness of the fear response is also demonstrated in animals; for example, laboratory-reared rats confronted with danger (e.g., a cat) for the very first time exhibit changes in autonomic nervous system activities and behavioral readiness (responses that are remarkably similar to those seen in humans), and ultrasonic (22 kHz) distress vocalization (a response that is an example of an SSDR) (e.g., Blanchard and Blanchard 1971, Blanchard et al. 1991, Bolles 1970).

Several lines of evidence point to temporal lobe structures (specifically the amygdala, an almond-shaped group of nuclei buried deep within the temporal lobes) as the key neural system underlying the central state of fear (see LeDoux 1996). The classic studies of Kluver and Bucy (e.g., 1937), who examined the effect of temporal lobe ablations in rhesus monkeys, were pioneering in this respect. Kluver and Bucy noted dramatic postlesion behavioral changes, including hyperactivity, hypersexuality, and a so-called ‘psychic blindness’ that was manifested as an apparent indifference to the significance of previously frightening and/or arousing stimuli. While the monkeys were wild and aggressive prior to surgery, they were remarkably tame and agreeable to human contact afterwards, suggesting that a crucial emotive brain center had been removed or significantly affected by the lesions. Subsequent work, such as that of MacLean and Delgado (1953) using focal stimulation of temporal lobe structures in awake, behaving monkeys, pointed to the amygdala as the critical site. The importance of the amygdala in fear has been recon- firmed in numerous studies employing diverse species of animals (e.g., rats, mice, cats, rabbits, monkeys) and various experimental techniques (e.g., lesions, stimulation, drug infusions, recordings), and most current theories of amygdalar function reflect the importance of the structure in the processing of emotional stimuli (see LeDoux 1996). Consistent with this view, recent human neuropsychological and brain-imaging studies have found that patients with amygdalar damage are selectively impaired in the recognition of facial expressions of fear (Adolphs et al. 1994), and that there is a significantly increased blood flow to the amygdala (as measured by functional magnetic resonance imaging, fMRI) when normal subjects are presented with pictures of fearful faces (Morris et al. 1996).
The amygdala, as one of the principal structures of the limbic system, has access to sensory input from diverse areas of the brain (e.g., olfactory cortex, hippocampus, thalamus, hypothalamus, neocortex), and projects to various autonomic and somatomotor structures that mediate specific fear responses (e.g., bed nucleus of stria terminalis for activating stress hormones, periaqueductal gray matter for defensive behavior, lateral hypothalamus for sympathetic activation) (see LeDoux 1996).
Given this pattern of connectivity, it is not surprising that damage to the amygdala eliminates stereotypical fear responses, such as defensive freezing (characterized as a motionless, crouching posture that functions to reduce the probability of detection by predators) exhibited by rodents in the presence of predators (Blanchard and Blanchard 1971). Nevertheless, while the amygdala is apparently an important component of the central fear-processing system, it does not necessarily follow that the amygdala is the seat of fear as an emotion (or a feeling). Given its intricate reciprocal connections with the neocortex, a structure implicated in higher cognitive functioning, it is very likely that the interactions between amygdala and neocortex ( particularly sensory and limbic association cortices) are responsible for the emotion of fear. Thus, although the amygdala is essential in activating various fear responses (or SSDRs) in both humans and rats, the fact that there are great differences in the neocortices of humans and rats suggest that perhaps there might be important differences in fear as experienced by the two species.
3. Acquired Fear
While humans and other animals are genetically predisposed to fear certain stimuli events, it is also adaptive for animals to have the capacity to learn about new dangers in their environments. For instance, although newborn infants innately exhibit fear to certain stimuli (e.g., loud noises), they do not show innate fear to flame or height (two stimuli that most children learn and adults remember to avoid). Thus, fear responses to many stimuli and events in the environment appear to be acquired.
Classical or Pavlovian fear conditioning has been widely employed for studying the mechanism(s) by which fear is acquired. Fear conditioning occurs when initially neutral conditioned stimuli (CSs) such as tones, lights, or distinctive environmental settings, are contingently paired with aversive unconditioned stimuli (USs) such as electric shocks, which reflexively activate unconditioned fear responses (URs). Through CS–US association formation, the CS comes to elicit various conditioned fear responses (CRs) that are similar to innate fear responses (Fig. 2, B). Perhaps the best known example of fear conditioning is the Little Albert experiment by Watson and Rayner (1920). Little Albert was an 11-month-old infant who initially exhibited curiosity (no fear) to a white rat by touching and playing with it. Watson and Rayner attempted to condition Albert to fear the rat by striking a steel bar with a hammer behind his head (US) as his hand touched the rat, causing him to startle, fall forward, and cry (UR). A week later, when the rat (CS) was placed near Little Albert’s hand, he withdrew his hand and began to cry (CR). This fear of the rat was also said to be observed in the presence of other white furry animals and objects (e.g., rabbits, dogs, fur muffs).
Modern investigations of fear conditioning typically employ rats as subjects and use a tone as a CS and a mild footshock as an US. Under these circumstances a single CS–US pairing results in robust (rapid and enduring) fear learning, as evidenced by a variety of conditioned fear responses exhibited upon subsequent presentations of the tone. Thus, the biological significance of fear conditioning is reflected by the quick and permanent nature of changes in the behavior toward the initially neutral stimulus that has been paired with an aversive experience.
Fear can also be rapidly acquired through instrumental or operant conditioning, in which the presentation of an aversive stimulus is contingent upon the behavior of the animal. A widely employed procedure with rodents is the passive (or inhibitory) avoidance task (see McGaugh et al. 1996). In this paradigm, the animal’s response (e.g., entering a dark compartment of a box when placed in an adjacent lighted compartment, or stepping down from a platform onto a grid floor) is paired with an aversive experience (e.g., a footshock). As a function of this response–stimulus pairing, the animal learns to avoid making the response that was followed by the aversive experience.
Similar to innate fear, evidence from lesion, pharmacological, recording, and stimulation studies indicates that the amygdala is essential not only to innate fear, but also to the acquisition of fear, as with fear conditioning (see Lavond et al. 1993, LeDoux 1996). Thus, amygdalar lesions ( particularly lesions of the central nucleus) impair the acquisition (learning) and retention ( performance) of various fear-related behaviors in rats, rabbits, and cats; neurons in the central nucleus of the amygdala are responsive to both conditioned and innate fear stimuli and undergo plastic changes during fear conditioning; and restricted stimulation of the amygdala elicits specific fear responses (e.g., freezing). Recent findings from human studies that patients with amygdalar damage are impaired in fear conditioning, and that blood flow to the amygdala is increased during fear conditioning in normal subjects, further add to the evidence that the amygdala is crucially involved in fear learning.
One hypothesis of amygdalar function in fear conditioning is presented schematically in Fig. 3. In this model, an emotional stimulus (such as a tone that has been paired with a foot-shock) is presented, and its sensory characteristics are processed by way of two parallel pathways that ultimately converge in the amygdala. One of these pathways is ‘direct’ and provides relatively ‘raw,’ unprocessed information about the stimulus, but does so very quickly. If the neocortex is the seat of consciousness, this thalamic pathway to the amygdala can be considered the subconscious (nondeclarative or implicit) fear route. The second pathway is ‘indirect’ and provides highly processed information about the stimulus, but does so at the cost of an increased processing time (LeDoux 1996).
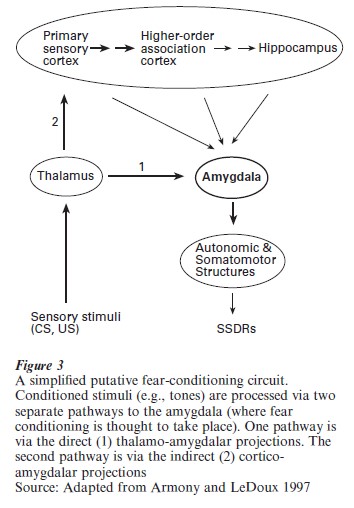
This cortical (conscious, declarative, or explicit) pathway consists of projections through primary auditory cortex to higher-order auditory association areas and the hippocampal formation, which then project to the amygdala. Upon reaching the amygdala (specifically the lateral basolateral nuclei), these two converging representations of the stimulus are subjected to additional processing within the amygdala itself. Most current theories assume that the essential plasticity underlying this form of learning occurs in the amygdala and corresponds to a neural instantiation of the association between the CS (the tone) and the US (the footshock) long assumed by psychologists to lie at the heart of classically conditioned behavior. The precise functions of the inputs to and outputs from the amygdala, as well as the intraamygdaloid processing of CS and US-related information, remain a topic of intensive research.
4. Fear Conditioning And Clinical Implications
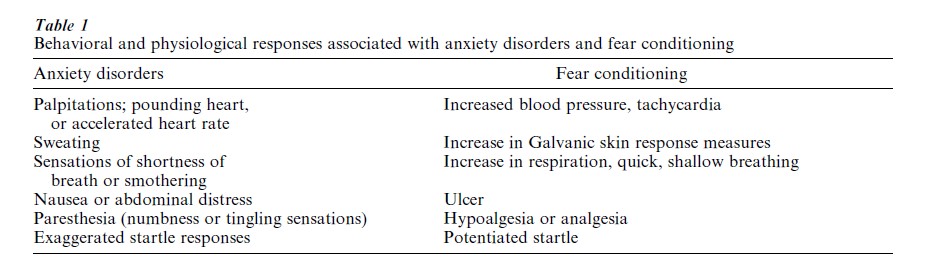
Fear conditioning serves as a model system for understanding not only the neural basis of acquired fear in nonhuman animals, but also the etiology of a variety of fear-related disorders in humans (e.g., anxiety, phobias, post-traumatic stress disorder, and panic disorder). Since the characteristic behavioral and physiological symptoms of human anxiety disorders closely resemble many of the fear responses observed in fear conditioning paradigms, it is possible that anxiety and fear are closely related (Table 1). (It is important to note, however, that the fact that the symptomatic manifestation of anxiety disorders is similar to the fear responses observed in fear conditioning does not necessarily mean that the underlying neuronal substrates of fear and anxiety are the same ( post hoc, ergo propter hoc).) While anxiety disorders can arise unpredictably, it is known that most patients experience their first symptoms in early adult life, indicating that maladaptive anxiety may be at least partly acquired. Indeed, clinicians from the time of Freud have speculated that anxiety patients have acquired their disorder as a result of traumatic learning experiences. Since the paradigm par excellence of traumatic learning is fear conditioning, fear conditioning appears to be useful in understanding the genesis of anxiety disorders (Fig. 3).
The question of whether anxiety is analogous to fear conditioning might be one of semantics; the difference is between normal, adaptive fear and abnormal, maladaptive fear (anxiety, phobias). However, there is an important distinction between anxiety disorders and fear conditioning (e.g., Seligman 1971) that might be taken to suggest that the two have very different bases. That is, while conditioned fear responses may be extinguished by, for example, actively preventing an animal from making escape or avoidance responses to a danger-signaling stimulus, fear-related disorders in humans (e.g., phobias) are quite resistant to extinction. In fact, anxiety disorders are characterized by fears that the patients recognize to be irrational but are unable to control.
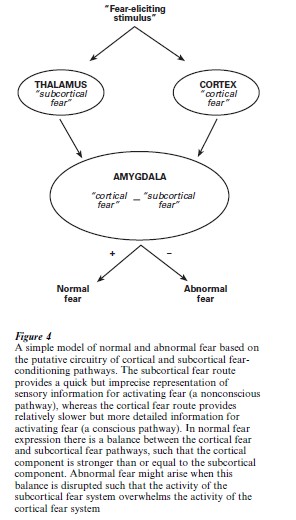
Based on Fig. 3, it is interesting to speculate that fear-related disorders (irrational fears) might be due to unbalanced activities in the cortical and thalamic pathways to the amygdala. For example, in most people a snake seen behind a glass window will elicit a different response than will a snake encountered in the woods. Both cortical and subcortical routes will process information about the snake, but the cortical (conscious) route will integrate the contextual information of the glass window and assess the safety of the situation ( possibly acting as a gate). However, if the cortical pathway is somehow disrupted, the subcortical activity will be left unchecked to activate fear, even in the inappropriate context. It is possible that this type of unregulated subconscious activation of fear contributes to the etiology of fear-related disorders.
Figure 4 illustrates a hypothetical model of normal, adaptive fear and abnormal maladaptive fear. It is conceivable that genetic predisposition and environmental factors (e.g., stress) interact in such a way as to disrupt the normal balance between cortically activated fear and subcortically activated fear, leading to fear-related disorders. If it is then supposed, as some have suggested (LeDoux 1996), that the direct thalamic inputs to the amygdala are relatively resistant to extinction, whereas the cortical inputs are not, it follows that fear disorders based on over-activity in the subcortical pathway should be ‘irrational,’ in the sense that they do not subside even when the patient recognizes them as such. A more comprehensive understanding of the neuronal substrates and, subsequently, the molecular mechanisms of the fear system will be useful in understanding and developing treatments for fear-related disorders.
5. Conclusion
The study of fear, a form of emotion that evolved relatively recently among the phylogeny of successful defensive mechanisms, is but one means of understanding emotion, but it is the most fruitful one to have been employed to date. This is likely due to the primacy of fear: unlike other emotions, fear is apparently shared by a wide variety of species. This ubiquity speaks to its importance as a mechanism for self-preservation.
References:
- Adolphs R, Tranel D, Damasio H, Damasio A 1994 Impaired recognition of emotion in facial expressions following bilateral damage to the human amygdala. Nature 372: 669–72
- Armony J L, LeDoux J E 1997 How the brain processes emotional information. Annals of the New York Academy of Science 821: 259–70
- Blanchard R J, Blanchard D C 1971 Defensive reactions in the albino rat. Learning and Motivation 2: 351–62
- Blanchard R J, Blanchard D C, Agullana R, Weiss S M 1991 Twenty-two kHz alarm cries to presentation of a predator, by laboratory rats living in visible burrow systems. Physiology and Behavior 50: 967–72
- Bolles R C 1970 Species-specific defensive reactions and avoidance learning. Psychological Review 71: 32–48
- Darwin C 1872 The Expression of the Emotions in Man and Animals. Murray, London
- Kluver H, Bucy P C 1937 ‘Psychic blindness’ and other symptoms following bilateral temporal lobectomy in rhesus monkeys. American Journal of Physiology—Regulatory Integrative and Comparative Physiology 119: 352–3
- Lavond D G, Kim J J, Thompson R F 1993 Mammalian brain substrates of aversive classical conditioning. Annual Review of Psychology 44: 317–42
- LeDoux J 1996 The Emotional Brain: the Mysterious Underpinning of Emotional Life. Simon and Schuster, New York
- MacLean P D, Delgado J M R 1953 Electrical and chemical stimulation of frontotemporal portion of limbic system in the waking animal. EEG Clinical Neurophysiology 5: 91–100
- McGaugh J L, Cahill L, Roozendaal B 1996 Involvement of the amygdala in memory storage: Interaction with other brain systems. Proceedings of the National Academy of Sciences of the United States of America 93: 13508–14
- Morris J S, Frith C D, Perrett D I, Rowland D, Young A W, Calder A J, Dolan R J 1996 A differential neural response in the human amygdala to fearful and happy facial expressions. Nature 383: 812–15
- Seligman M E P 1971 Phobias and preparedness. Behavioral Therapy 2: 307–20
- Watson J B 1924 Behaviorism. Norton, New York
- Watson J B, Rayner R 1920 Conditioned emotional reactions. Journal of Experimental Psychology—Human Perception and Performance 3: 1–14
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality