
Sample Comparative Psychology Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. iResearchNet offers academic assignment help for students all over the world: writing from scratch, editing, proofreading, problem solving, from essays to dissertations, from humanities to STEM. We offer full confidentiality, safe payment, originality, and money-back guarantee. Secure your academic success with our risk-free services.
Comparative psychology is a diverse field, one which is characterized by at least four different approaches to its subject matter. The most venerable of these approaches is to describe similarities and differences in the behavioral and psychological abilities or capacities of various species, irrespective of their evolutionary lineage (relationship) (see Sect. 1). A variant of this first approach is to study the behavior or learning abilities of various species with a view toward describing the advancement or progression of psychological functioning going from the more simple to the more complex, in lines that are not necessarily closely related in an evolutionary sense; this approach is called anagenesis (or the levels approach) and typically shows improvements in developmental plasticity, learning ability, etc. in more recently evolved forms. The point of this approach is to discern broad trends in evolution, especially psychological evolution, and it is based on a hierarchical conception of learning abilities, going from the simple to the complex.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
A more recent variant of that approach is to describe similarities and differences in species that are closely related in an evolutionary sense, and to try to interpret these results with respect to the evolution of particular behavior (see Sect. 2). Another approach practiced by comparative psychologists is to study the species-typical behavior of animals in the field throughout their life cycle (see Sect. 3). That approach was made more popular by the zoologically-trained animal behaviorists in Europe, who called themselves ethologists to distinguish themselves from psychologically-trained animal behaviorists in the United States, many but not all of whom confined their studies to the laboratory. Finally, there is the animal model approach, which tries to find nonhuman species with behavioral or psychological repertoires that are similar to humans so that the results of experiments with the model may throw light on seemingly related behavior in human beings (see Sect. 4).
As stated by T. C. Schneirla (1962, p. 690R), ‘Comparative psychology is studied because the behaviour of animals is in itself interesting, because of important applications in fields such as medicine and animal training, and also because of significant bearings on human psychology.’ The remainder of this research paper gives examples of each of these approaches.
1. Similarities And Differences In Distantly Related Species
One of the most fundamental similarities in animals of all species is their ability to stop responding to a repeated stimulus in the absence of reward. This phenomenon is called habituation and it is, along with sensitization (described next), the lowest form of learning. It is of course adaptive for animals to cease responding to a stimulus event that poses no threat and offers no reward. In snails, for example, when the substrate upon which they are walking is given a mechanical shock, they withdraw their tentacles for a brief period. When the shock is repeated in exactly the same way at regular intervals, the extent and duration of tentacle withdrawal steadily decreases until finally the animal becomes completely indifferent to the stimulus. Sensitization is the obverse of habituation in that animals will orient to abruptly presented sights, sounds, smell, etc., and their orientation reaction will be quicker and longer if the stimulus changes somewhat from trial to trial. Both habituation and sensitization are nicely demonstrated in an experiment using the wiping reflex of frogs. When the frogs are touched with a hair on precisely the same zone of skin from trial to trial, their wiping reflex declines (fewer wipes per 100 stimuli). However, when the same hair is used to stimulate the frog’s skin at somewhat different places each time, the wiping reflex is enhanced (more wipes per 100 stimuli). The next higher form of learning is called associative (stimulus– response or S–R) learning or conditioning, and was made popular by Ivan Pavlov’s experiments with the salivary response of dogs. In these experiments, a bell or tone was sounded followed by the sight of meat powder. The hungry dog salivated at the sight and smell of the meat powder (unconditioned response). After numerous pairings of the to-be-conditioned stimulus (tone) with the unconditioned stimulus (meat powder), the dog began to salivate at the sound of the tone (conditioned response) in advance of the presence of the meat powder. Associative learning of this kind—called classical conditioning—has been demonstrated in all vertebrates from fish (‘lowest’ vertebrate) to humans (‘highest’ vertebrate). It has also been demonstrated in some invertebrate species, but not in all, so this must be considered a higher level of learning than habituation and sensitization. There is another form of conditioning—operant learning—that strengthens existing responses through reward (food for a hungry animal, e.g.) or reduces existing responses through punishment (electrical shock, e.g.). This form of conditioning is widely practiced among animal trainers and all vertebrates and some invertebrates are capable of it. The next psychological level as far as learning is concerned is trial-and-error or selective learning, in which there is a significant perceptual, perhaps cognitive, component, in the sense that the animal must discover a specific relationship between its activities and a goal object. As first studied by E. L. Thorndike in the late 1890s in connection with his doctoral dissertation, a variety of vertebrate species were placed in a puzzle box or a maze, from which they had to escape in order to get food or join companions. Thorndike’s idea was that the animal had to learn to associate making certain movements (R) in a problem situation(s) (pulling a cord or unlatching a lock) with a subsequent ‘satisfying’ state of affairs (getting to food or joining companions). So the S–R association in this context is the situation and the successful movement leading to the satisfying state of affairs. While some invertebrate species can do trial and error learning, this class of learning reaches its peak in mammals. The highest category of learning ability, considered by some the true criterion of intelligent behavior, is reasoning or, in other terms, insightful problem-solving. This is seen in nonhuman animals when an organism solves a problem by using as a tool (means) some object(s) that never previously in that animal’s experience had been used in that way. The most famous instance of this kind was Wolfgang Kohler’s chimpanzee Sultan, who, in order to reach a banana high in his cage, stacked boxes one upon another to make a ladder enabling him to reach the banana. While no doubt occurring in humans and chimpanzees, comparative psychologists have been sparing in applying this appelation to other mam malian species.
There is a form of learning that takes place through mere exposure to physical and social objects; the exposure leads to subsequent attachment, preference, or acceptance of these objects (i.e., animals and humans not exposed to these objects do not show attachment, preferences, or acceptance of these objects). This familiarity mechanism is pervasive; it is found in many invertebrate species as well as vertebrates (Szentesi and Jermy 1989), and it is often referred to as imprinting, although, technically speaking, imprinting involves a sensitive period early in life and is usually defined as inducing an enduring preference not merely acceptance (nonavoidance) of an object. While particularly striking in young animals (Sluckin 1965), exposure learning operates throughout life in some species such as our own (Hebb 1946, Zajonc 1971). Exposure learning is difficult to classify with certainty in the hierarchical scheme adopted here. Since it is so pervasive and is brought to bear by mere exposure, it probably fits in above the level of habituation and sensitization because of its requirement for a higher degree of perceptual differentiation and longer term memory. It would almost always be adaptive to form attachments, preferences, and acceptance of familiar physical and social objects.
There are two approaches to classification: lumping and splitting. The hierarchical classification of adaptive or adjustive learning abilities into habituation, sensitization, exposure learning, conditioning, trial-and-error learning, and reasoning is an example of lumping. In 1971, Gregory Razran published a book, Mind in Evolution: An East–West Synthesis of Learned Behavior and Cognition, in which he described 11 forms or levels of learning. He included the ones described here (except exposure learning) and further broke down the various forms of conditioning and also added ‘symboling’ to include the role of language in what Razran called ‘the learning of thinking, planning, and willing.’ Razran’s treatise is without equal in the literature concerning the comparative analysis of learning ability in a large number of species, as well as introducing Western scientists to the large literature that appears only in the Russian language.
A striking example of behavioral anagenesis is provided by an experiment in which the progression in learning a maze is observed in ants and rats (Schneirla 1962). As shown in Fig. 1, the ants took more than twice the number of trials to get their error rate down to near zero, as compared with the rats. Not only do the rats learn the maze much more quickly than the ants, they do it in a different fashion. Note the large reductions in errors from trial to trial in the rats’ learning curve compared with the ants’, in which there is slow and gradual progress.
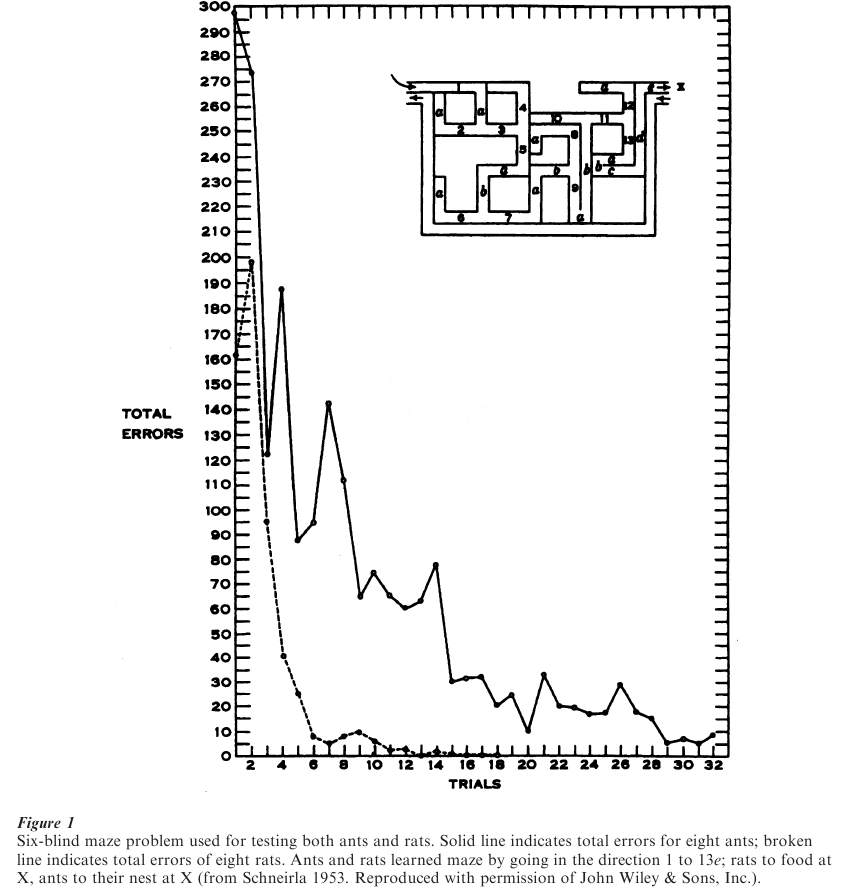
Schneirla observed not only a quantitative difference in the behavior of the rats and ants but also a qualitative difference:
Not only do the rats make fewer errors and require fewer runs than the ants to master this maze problem, but their learning process in it differs markedly in several respects from that of the ants. The three stages so clearly marked in the ants cannot be distinguished reliably in the rats. The rats, in fact, progress from the very first run in shortening and eliminating their entrances into the blind alleys; moreover they drop out some of the blinds quickly and almost as units and others ‘by jumps’ although less rapidly. The ant’s pattern of eliminating blind-alley entrances gradually by small degrees is very unlike the rat’s response to this problem (Schneirla 1962, p. 513).
Anagenic trends in learning (improvements over the course of evolution) have always been considered in light of evolutionary changes in brain structure and function. In an instructive experiment with three species of birds and three species of mammals, L. V. Krushinsky and colleagues (1965) examined what he considered the ability of these different species to solve an ‘elementary logical problem’ by what he called ‘extrapolation’ without any specific training. The task involved the animals facing a screen with a gap in its center. A bowl of food passed the gap moving either in the right or left direction, remaining out of sight behind the screen once having passed the gap. The animal was free to move behind either end of the screen during one minute after the bowl passed the gap. The question was whether the animal would walk around the screen on the side of the food bowl. The number of trials each animal responded and the side to which they responded was recorded. Each animal was tested two times within one week and were given 4–8 trials during each test, with the food bowl moving alternately in both directions from trial to trial.
As can be seen in Table 1, among the three bird species, the pigeons responded only twice in 27 trials, the chickens (hens) responded in slightly over one-half of the trials but their choice of sides was at a chance level, and the crows responded at a high rate (85.7 percent) and chose the correct side 71.4 percent of the time. Among the three mammalian species, the rabbits responded on only 27 percent of the trials (9) but they were correct on eight of the nine trials. The cats showed a high level of response but their choice was at a chance level (21 times to the side with food and 15 times to the empty side). The dogs responded on almost 90 percent of the trials and their correct performance (77.8 percent) was somewhat better than the crows (71.4 percent).
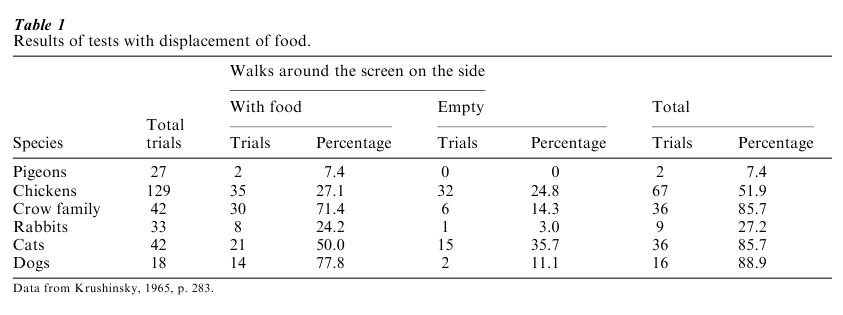
Psychologically speaking, Krushinsky’s task involves at least three components: object permanence (the animal must be aware that an object still exists even though it has passed from view), logical inference (it must be on this side rather than that side, depending on the bowl’s direction of movement), and detour learning (the animal must be able to move around an obstacle without being able to see the food). There is a great deal of variation among avian species in a structure of the brain called the wulst. The high level of performance of the birds in the crow family was attributed by Krushinsky to the high level of differentiation of their wulst in comparison to chickens and pigeons, the latter having the least developed wulst. The wulst is believed to be homologous (i.e., it is the ‘same’ organ in a different species) to the neocortex in mammals. Given the poor performance of the rabbits, if Krushinsky’s reasoning is correct, one would predict a low level of neocortical development in rabbits compared with cats and dogs, and, to carry the comparison one step further, a difference between cats and dogs in favor of dogs. It is not known whether these predictions are correct or not.
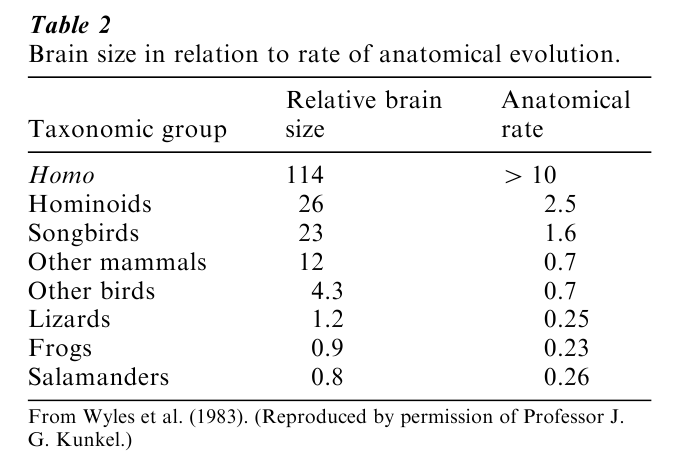
The anagenesis of the brain that does not have precise parallels in psychological function and behavior is the increase in the ratio of brain size to body size that has occurred in the vertebrate phylum (fish, amphibians, reptiles, birds, and mammals). As shown in Fig. 2, the size of the brain of birds and mammals exceeds that of all other vertebrates at all body sizes. This rather gross statistic would suggest important psychological differences between birds and mammals and the other three classes of vertebrates, and has given some validity to the division of the phylum into ‘higher’ (birds, mammals) and ‘lower’ (reptiles, amphibians, fish) vertebrates. In the terms already developed in this research paper, one should expect to see, by and large, greater behavioral plasticity in birds and mammals than in lower vertebrates. As it happens, that pre-diction does accord rather well with learning ability, especially of a species’ ability to show forms of learning above the level of conditioning: sensory preconditioning and learned stimulus configuring are possible only in birds and mammals, according to Razran’s (1971) extensive review of the literature. In terms of exploratory behavior in the face of novel objects, in a study of over 300 animals in over 100 different species, there was a consistent finding of mammalian superiority over the other vertebrate species (Glickman and Sroges 1966). Finally, given the presumptive relationship between large brains and behavioral plasticity, species with large brains showing a greater degree of exploratory behavior, may be evidence of a faster evolutionary pace in these species than in species with smaller brains. That expectation accords rather well with the findings of Wyles et al. (1983) of an almost perfect correlation between relative brain size and rate of anatomical evolution for a large number of vertebrate species.
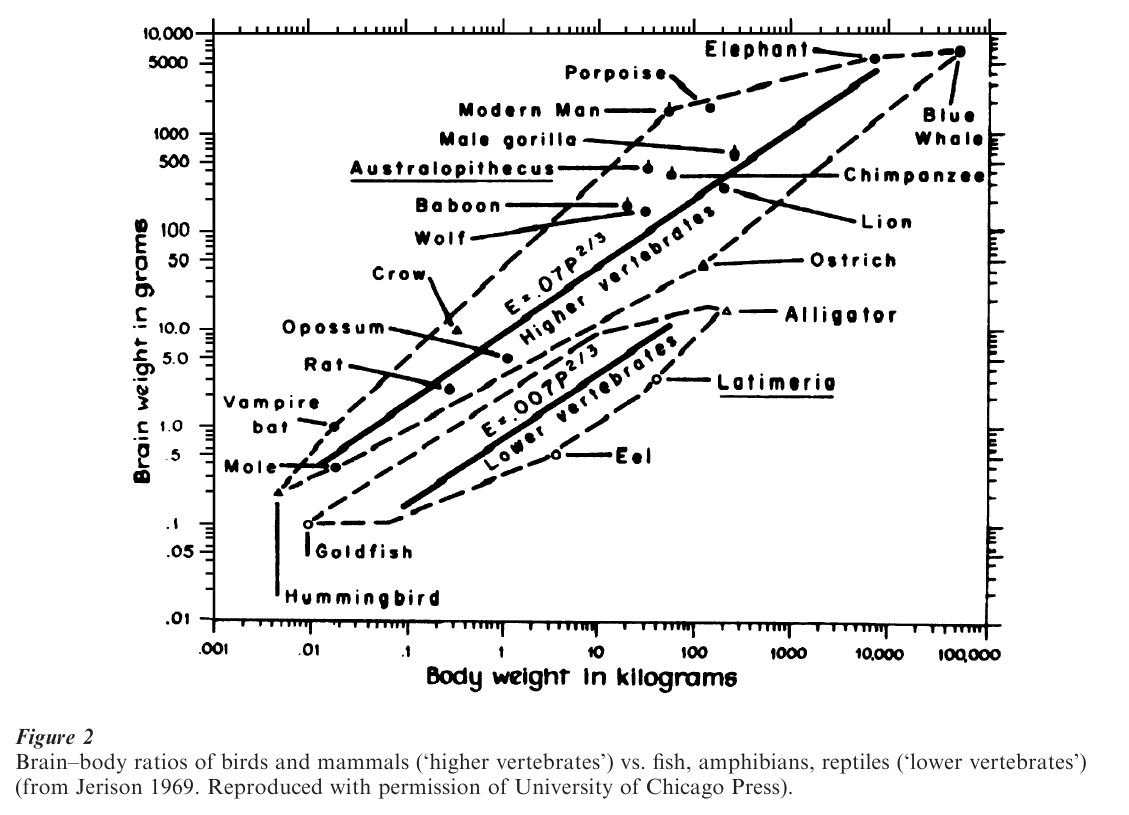
As shown in Table 2, humans, the group with the largest relative brain size (Fig. 2), show the fastest rate of anatomical evolution, with the larger and older hominoid groups (human-like forms) ranking second in brain size and rate of evolution. What is perhaps most interesting is that the relatively recently evolved songbirds rank just below hominoids and well above other mammals and other birds on both relative brain size and rate of anatomical evolution. Finally, the classifications of ‘other mammals’ and ‘other birds’ show a relatively larger brain size than the lower vertebrates (lizards, frogs, salamanders) and a corresponding faster tempo of evolutionary change. Variations in early experience during the course of individual development also contribute in an essential way to exploratory behavior and learning ability in adulthood (Gottlieb 1992), so it is not solely the possession of a large brain that is responsible for these outcomes.
2. Similarities And Differences In Closely Related Species: Importance Of Early Experience And Context
Humans share psychological and social characteristics with other primates. Language has always been considered the greatest difference between humans and other primates. Nonhuman primates do not share our vocal apparatus and thus cannot speak, so, in order to study linguistic competence in our relatives, manual sign language or tokens have been used. There are three components to language: syntax (grammar), semantics (meaning), and pragmatics (using language to attain goals). There is little doubt that chimpanzees in particular show evidence of the latter two and some primitive understanding of grammar (e.g., changing word order to alter the meaning of a phrase or sentence). What has hampered a genuine testing of the limits of chimpanzees’ and other nonhuman primates’ linguistic competence has been the age of the subjects when language instruction was started. Adults are inferior in language learning to juveniles, and it is only quite recently that a bonobo (pygmy chimpanzee) has been started in infancy and the as yet unpublished results are said to represent quite an improvement over previous attempts (Savage-Rumbaugh et al. 1998).
The importance of the presence and absence of early learning or experiential opportunities has not always been appreciated in comparative psychology, although the otherwise influential scientist T. C. Schneirla advocated the study of individual development as ‘the backbone of comparative psychology.’ An example of the influence of different developmental opportunities for learning is the use of pointing in chimpanzees as a communicative gesture to indicate an object of interest to a conspecific partner or human caretaker. Language-trained chimpanzees point with their index finger, as do humans aged 13–18 months. Language- naive chimpanzees point with their whole hand, as do humans aged 8–12 months (Leavens and Hopkins 1999). So developmental experience with linguistic training fosters pointing with the index finger in nonhuman primates and it parallels the beginning use of words in the older human infant (pointing frequently accompanies naming in human infants and their primary caretakers).
The importance of development and rearing context in comparing closely related species is shown in studies of domesticated animals compared with their wild (free-living) ancestors. Such studies uniformly show a reduction of aggressiveness, an increase in social tolerance, and a decrease in the domestic organism’s physiological stress axes (Kunzl and Sachser 1999). These are inevitable accommodations to human-made housing and living conditions. The importance of environmental context is shown in a study by D. B. Miller (1977) in which he compared the courtship patterns of domestic and wild mallard ducks. In the barnyard situation, domestic mallards do not show courtship patterns and it was believed that these patterns were lost due to ‘degenerate’ genetic changes presumed to be inherent in the domestication process. Miller discovered that, when placed on a farm pond, domestic mallard not only showed all the courtship patterns of their wild progenitors but also showed them in the same order: the courtship and mating patterns were undistinguishable between the wild and domestic birds when the more natural species-typical context was provided.
3. The Importance Of Field Studies Of Animals Living In Their Usual Habitat
While some comparative psychologists have done (and do) field studies, most comparative psychology was (and is) studied in the laboratory. Laboratory analysis is more precisely controllable than fieldwork, and so it is a necessary adjunct to field observations. For a period of time in the mid-twentieth century, much of comparative psychology lost touch with natural eco-logical settings and, consequently, the relevance of the laboratory findings to a species’ adaptation in its usual abode became dubious. (There was also an increased tendency to study a narrow range of readily available domesticated species on a narrow range of problems dictated by general theories of behavior that held sway for too many years in experimental psychology.) It is largely to the credit of the zoologically trained European thologists that ecological relevance and the study of animals as they exist in nature became focal points. The observations in naturalistic settings pro-vide the problems (questions) to be answered by experiments in the laboratory, so the results of the laboratory experiments are ecologically valid.
The study of imprinting in waterfowl and other precocial bird species is a case in point. Imprinting was believed to be the process whereby a young bird learned its species identification. It was thought that the young birds’ perceptual system was wide open (like a blank slate), so all sorts of nonspecies-typical artifacts were used in the laboratory as imprinting objects, because it was convenient and the young incubator-hatched birds would indeed approach, follow, and become attached to these visual objects. Field observations (by a comparative psychologist!) indicated that, in a number of species, the mother uttered a special call to which the young were highly attracted. As subsequently shown in laboratory experiments, even incubator-hatched ducklings and chicks were attracted to the maternal call of their species, even though they had not previously heard it, having been hatched in an incubator (Gottlieb 1971). Those field observations, and the subsequent laboratory experiments, cast serious doubt on the ecological validity of the prior experiments on visual imprinting, especially when it was shown in the laboratory that the incubator-hatched birds were more attracted to the maternal call than to the visual aspect of a hen of their species. Because of their attraction to the maternal call, in nature young birds would always follow a hen of their own species, not just any object that happened to be passing by as implied by laboratory studies of visual imprinting.
4. Animal Models
The question of the utility of animal models for understanding human psychological and behavioral development does not seem tenable, if by animal model we mean we are modeling identical or even highly similar human psychological, social, and behavioral phenomena. On the other hand, the validity of animal models of human genetic, physiological, and anatomical outcomes seems much more likely because the analytic techniques are available to specify the identity of the genes, physiology, and anatomy in question. In that case, it is much easier to know whether one is right or wrong about the model. The certainty that animal models are faithfully mimicking their presumed human counterparts in the arena of psychological, social, and behavioral functions is always open to question. On the other hand, the outcome of programmatic developmental experiments with animals can sometimes provide interesting and significant food for thought—hypotheses not facts— about human development. Also, insofar as they do that they can also provide food for thought about development in other animal species. So, it is at the level of generalities, not specifics, that animal experiments are useful for thinking about human development.
Animal models may not do justice to human development when they emphasize (or seem to implicate) single developmental pathways to psycho-logical or behavioral outcomes. Human development involves equifinality (more than one pathway to a given outcome), a principle of extreme importance, especially when dealing with problem populations (psychological disorders): there is likely not only one way to intervene (or develop the disorder in the first place).
An excellent example of how comparisons of animal models and human psychological functioning can mutually enrich each other is provided by experimental studies of prenatal and early postnatal experience in the development of intersensory perception in human and animal infants (Lickliter and Bahrick 2000). By taking a comparative approach, these authors were able to show how prenatal factors, brain–behavior relations, and context and experience contribute to infants’ intersensory responsiveness. Another area in which human and animal comparisons has been mutually rewarding is the study of the role of early auditory experience in vocal development in young songbirds and human infants (Heaton and Brauth 1999).
Bibliography:
- Glickman S E, Sroges R W 1966 Curiosity in zoo animals. Behaviour 26: 151–88
- Gottlieb G 1971 Development of Species Identification in Birds: An Inquiry into the Prenatal Determinants of Perception. University of Chicago Press, Chicago
- Gottlieb G 1992 Individual Development and Evolution: The Genesis of Novel Behavior. Oxford University Press, New York
- Heaton J T, Brauth S E 1999 Effects of deafening on the development of nestling and juvenile vocalizations in budgerigars (Melopsittacus undulatus). Journal of Comparative Psychology 113: 314–20
- Hebb D O 1946 On the nature of fear. Psychological Review 53: 250–75
- Jerison H J 1969 Brain evolution and dinosaur brains. American Naturalist 103: 575–88
- Krushinsky L V 1965 Solution of elementary logical problems by animals on the basis of extrapolation. Progress in Brain Research 17: 280–308
- Kunzl C, Sachser N 1999 The behavioral endocrinology of domestication: A comparison between the domestic guinea pig (Ca ia aperea f. porcellus) and its wild ancestor, the cavy (Ca ia aperea). Hormonal Behavior 35: 28–37
- Leavens D A, Hopkins W D 1999 The whole-hand point: The structure and function of pointing from a comparative perspective. Journal of Comparative Psychology 113: 417–25
- Lickliter R, Bahrick L E 2000 The development of infant intersensory perception: Advantages of a comparative convergent-operations approach. Psychological Bulletin 126: 260–80
- Miller D B 1977 Social displays of mallard ducks (Anas platyrhynchos): Effects of domestication. Journal of Comparative and Physiological Psychology 91: 221–32
- Razran G 1971 Mind in Evolution: An East–West Synthesis of Learned Behavior and Cognition. Houghton Mifflin, Boston
- Savage-Rumbaugh S E, Shanker S G, Taylor T J 1998 Apes, Language, and the Human Mind. Oxford University Press, New York
- Schneirla T C 1953 Modifiability in insect behavior. In: Roeder K D (ed.) Insect Physiology. Wiley, New York
- Schneirla T C 1962 Psychology, comparative. In: Encyclopaedia Britannica, 690Q–703
- Sluckin W 1965 Imprinting and Early Learning. Aldine, Chicago
- Szentisi A, Jermy T 1989 The role of experience in host plant choice by phytophagous insects. In: Bernays E A (ed.) Insect–Plant Interactions. CRC Press, Boca Raton FL, Vol. 2, pp. 39–74
- Wyles J S, Kunkel J G, Wilson A C 1983 Birds, behavior, and anatomical evolution. Proceedings of the US National Academy of Sciences 80: 4394–7
- Zajonc R B 1971 Attraction, affiliation, and attachment. In: Eisenberg J F, Dillon W S (eds.) Man and Beast: Comparative Social Behavior. Smithsonian Institution Press, Washington, DC, pp. 143–79
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality