
Sample Learning By Occasion Setting Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
1. Simple Conditioning And Occasion Setting
In classical conditioning, organisms learn to predict the presence or the absence of an unconditioned stimulus (US) upon presentation of a conditioned stimulus (CS). Whereas prediction of the presence of the US seems to be mediated by excitatory CS–US associations, prediction of the absence of the US seems to be mediated by inhibitory CS–US associations. When the CS predicts that the US will be present, a conditioned response (CR) is generated. When several CSs are presented together they can combine their predictions and determine whether the CR is generated.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
Although CSs are generally considered to only act in the role of ‘simple CS’ as described in the previous paragraph, experimental data show that a CS can also play as an ‘occasion setter.’ Whereas as a simple CS a CS forms excitatory or inhibitory associations with a US, as an occasion setter, it modulates the CR generated by another CS.
These alternative roles played by a CS were established in experiments that included different temporal arrangements of feature positive (FP) and feature negative (FN) discriminations. In a FP discrimination, the ‘feature’ (X) and the ‘target’ (A) are presented together with the US, and these presentations alternate with presentations of A alone. In a FN discrimination, XA compounds are presented in the absence of the US while A is presented with the US. FP and FN discriminations are simultaneous when X and A are presented at the same time, and serial when the presentation of X precedes the presentation of A.
Using simultaneous and serial FP and FN discriminations, Holland (1983, 1992) offered the following three criteria to distinguish simple conditioning from occasion setting roles of a CS.
1.1 Response Form
Under food deprivation, the form of the CR evoked by a light and tone compound reveals its associative origins: if the CR is rear behavior, then responding is the consequence of light-food associations, but if the CR is head jerk behavior, responding reflects tone-food associations. Taking advantage of this fact, Ross and Holland (1981) reported that in the simultaneous FP, the form of the CR during XA presentations was characteristic of X–US associations, whereas in the serial FP, the rats displayed CRs characteristic of A–US associations. Thus, Ross and Holland’s observations suggested that in a simultaneous FP discrimination X acts as a simple CS, but with a serial FP discrimination X acts as an occasion setter. A more complicated pattern of results for simultaneous FP discriminations as a function of the relative intensity of the feature and target stimuli was later reported by Holland (1989c).
1.2 Extinction And Counter-Conditioning
If performance on FP and FN discriminations is based on simple associations between X and the US, then elimination of those associations after discrimination training should abolish discriminative performance. Therefore, to the extent that rats solve FP discriminations by forming simple excitatory associations between X and the US, then repeated presentation of X alone after training should extinguish those associations, and consequently eliminate responding to the XA compound. Similarly, to the extent that rats solve FN discriminations by forming simple inhibitory associations between X and the US, then repeated reinforced presentations of X after training should establish excitatory X–US associations, and hence eliminate X’s simple inhibitory powers. Accordingly, in simultaneous FP and FN discriminations, Holland (1989b, 1989d) found that post-training extinction of X produced severe decrements in performance on FP discriminations, and post-training counterconditioning of X abolishes performance on FN discriminations. In contrast, in serial FP and FN discriminations, thought to establish occasion setting, post-training extinction or counter-conditioning of X had minimal effects (Holland 1989b, 1989d, Rescorla 1985, 1986).
1.3 Transfer
The result of combining simple CSs and occasion setters with other CSs has been studied in transfer experiments. For example, if a feature X previously trained in a simultaneous FP discrimination is paired with a trained-then-extinguished CS, B, the simple CS– US associations of the feature will immediately generate responses on XB trials (Holland 1986a, 1986b, Holland and Lamarre 1984). By contrast, if a feature X previously trained in a serial FP discrimination is paired with a trained-then-extinguished CS, B, the occasion-setting properties of the feature might (Davidson and Rescorla 1986, Jarrard and Davidson 1991) or might not generate responses on XB trials (Holland 1986a).
2. A Real-Time Model Of Pavlovian Conditioning
The alternative roles played by a CS can be best understood when described in terms of a formal model of classical conditioning presented by Schmajuk and DiCarlo (1992) and Schmajuk et al. (1998). The model is a real-time neural network in which CS inputs are connected to different CR outputs both directly and indirectly through a hidden-unit layer that codes configural stimuli. In the network, a CS acts as a simple stimulus through its direct connections with the output units and an occasion setter through its indirect configural connections via the hidden units. Schmajuk et al. (1998) showed through computer simulations that the network accounts for a large part of the data on occasion setting. Two examples for FP discriminations are shown in Sects. 2.1 and 2.2, (a) the conditions in which CS behaves as an occasion setter, and (b) how occasion setters are impervious to extinction.
2.1 Conditions In Which A CS Behaves As An Occasion Setter
The model explains why a CS behaves (a) as a simple CS in a simultaneous FP discrimination; or (b) as an occasion setter in a serial FP discrimination.
Simultaneous FP discrimination in Fig. 1 (top panels) illustrates the real-time values of X, A, and the US during reinforced and non-reinforced trials in a simultaneous FP discrimination. Figure 1 (bottom panel) summarizes the model’s solution for a simultaneous FP discrimination. Whereas X acts as a simple CS because it acquires a strong direct association with the US, A acquires a very weak association. Because direct associations of A and X with the US determine the response form, the model generates mostly CRs characteristic of X–US associations during compound XA presentations.
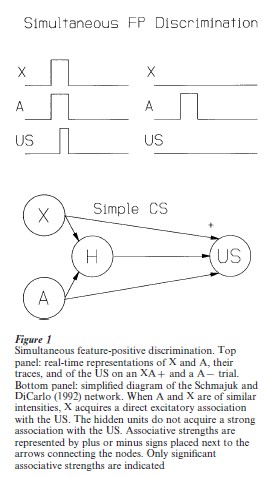
Serial FP discrimination in Fig. 2 (top panels) illustrates the real-time values of stimuli during reinforced trials in a serial FP discrimination. Figure 2 (bottom panel) shows a simplified depiction of the associative structure established in the network after serial discrimination trials. Because of their different temporal relations with the US, A accrues strong A–US associations but X only accrues weak X–US associations. Since the direct associations of X and A with the US determine the form of responding, responding on XA presentations comprises mostly CRs characteristic of A–US associations. Furthermore, responding on A-alone trials is small because it is strongly inhibited by the hidden units (which are activated by A), and responding on XA presentations is large because the inhibitory influence of the hidden units is ameliorated by the inhibitory action of X on the hidden units.
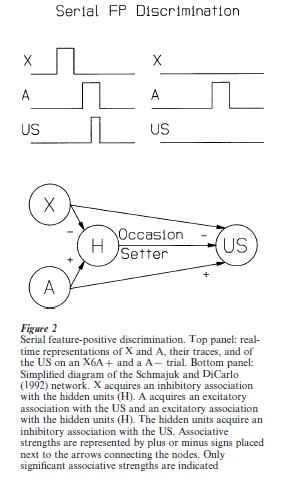
2.2 Occasion Setters Are Impervious To Extinction
Since simultaneous FP discriminations are solved by the model almost completely by direct X–US associations (see Fig. 1), the effect of X extinction is dramatic, abolishing CR responding on both X and XA test trials. By contrast, compound responding for serial FP is controlled by A–US associations which are attenuated by the inhibitory H–US associations (see Fig. 2). Since X–H and H–US associations are only minimally affected by X extinction, very little decrement is observed in XA compound responding after extinction.
3. Conclusion
A CS can behave either as a simple CS or as an occasion setter. In the first case, the CS acquires excitatory or inhibitory associations with the US, in the second case the CS might enhance or depress the effect of the simple association accrued by another CS. The properties of simple CSs and occasion setters have been thoroughly studied (for a review see Swartzentruber 1995).
A neural network model presented by Schmajuk et al. (1998) can accurately describe most of the available data. Other learning models that also include direct and indirect associations between CS inputs and CR outputs (e.g., Bouton 1994) also provide correct depictions of the phenomena. Neural networks without direct connections (e.g., Gluck and Myers 1993, Wagner 1992) can only describe some of the experimental results. Some of the physiological mechanisms of occasion setting have been addressed in terms of neural networks (Gluck and Myers 1994, Schmajuk and Buhusi 1997). A recent volume, published by Schmajuk and Holland (1998) presents the latest data and theoretical approaches to the subject.
Bibliography:
- Bouton M E 1994 Conditioning, remembering, and forgetting. Journal of Experimental Psychology: Animal Behavior Processes 20: 219–31
- Davidson T L, Rescorla R A 1986 Transfer of facilitation in the rat. Animal Learning and Behavior 14: 380–6
- Gluck M A, Myers C E 1993 Hippocampal mediation of stimulus representation: A computational theory. Hippocampus 3: 491–516
- Gluck M A, Myers C E 1994 Context, conditioning, and hippocampal representation in animal learning. Behavioral Neuroscience 108: 835–47
- Holland P C 1983 Occasion-setting in Pavlovian feature positive discriminations. In: Commons M L, Herrnstein R J, Wagner A R (eds.) Quantitative Analyses of Behavior: Discrimination Processes. Ballinger, New York, Vol. 4, pp. 183–206
- Holland P C 1986a Temporal determinants of occasion setting in feature positive discriminations. Animal Learning and Behavior 14: 111–20
- Holland P C 1986b Transfer after serial feature positive dis-crimination training. Learning and Motivation 17: 243–68
- Holland P C 1989a Acquisition and transfer of conditional discrimination performance. Journal of Experimental Psychology: Animal Behavior Processes 15: 154–65
- Holland P C 1989b Feature extinction enhances transfer of occasion setting. Animal Learning and Behavior 17: 269–79
- Holland P C 1989c Occasion setting with simultaneous com- pounds in rats. Journal of Experimental Psychology: Animal Behavior Processes 15: 183–93
- Holland P C 1989d Transfer of negative occasion setting and conditioned inhibition across conditioned and unconditioned stimuli. Journal of Experimental Psychology: Animal Behavior Processes 15: 311–28
- Holland P C 1992 Occasion setting in Pavlovian conditioning. In: Medin D L (ed.) The Psychology of Learning and Motivation. Academic Press, New York, Vol. 28, pp. 69–125
- Holland P C, Lamarre J 1984 Transfer of inhibition after serial and simultaneous feature negative discrimination training. Learning and Motivation 15: 219–43
- Jarrard L E, Davidson T L 1991 On the hippocampus and learned conditional responding: Effects of aspiration versus ibotenate lesions. Hippocampus 1: 103–13
- Rescorla R A 1985 Conditioned inhibition and facilitation. In: Miller R R, Spear N E (eds.) Information Processing in Animals: Conditioned Inhibition. Erlbaum, Hillsdale, NJ, pp. 299–326
- Rescorla R A 1986 Facilitation and excitation. Journal of Experimental Psychology: Animal Behavior Processes 12: 325–32
- Ross R T, Holland P C 1981 Conditioning of simultaneous and serial feature-positive discriminations. Animal Learning and Behavior 9: 293–303
- Schmajuk N A, Buhusi C 1997 Stimulus configuration, occasion setting, and the hippocampus. Behavioral Neuroscience 111: 235–58
- Schmajuk N A, DiCarlo J J 1992 Stimulus configuration, classical conditioning, and the hippocampus. Psychological Review 99: 268–305
- Schmajuk N A, Holland P C 1998 Occasion setting. In: Associative Learning and Cognition in Animals. American Psychological Association, Washington, DC
- Schmajuk N A, Lamoureux J, Holland P C 1998 Occasion setting and stimulus configuration: A neural network approach. Psychological Review 105: 3–32
- Swartzentruber D 1995 Modulatory mechanisms in Pavlovian conditioning. Animal Learning and Behavior 23: 123–43
- Wagner A R 1992 Some complexities anticipated by AESOP and other dual-representation theories. Paper abstracted in Kimmel H (Chair), Symposium on Pavlovian Conditioning with Complex Stimuli, XXV International Congress of Psychology. International Journal of Psychology: 101–2
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality