
Sample Sensory and Working Memory Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
To remember is to conjure up an image of the distant past, or perhaps a reflection from hours or days previous. Yet we remember over the very short term as well—over time periods lasting minutes, seconds, and even milliseconds. Consider language: We need to remember the early parts of a spoken phrase, or the particular sequence of phonemes in a word, for periods lasting beyond their physical presentation. Such short-lived memories are widely believed to be adaptive components of on-line cognitive processing. They help us produce and interpret spoken language, remember telephone numbers, reason, solve problems, and even think. The purpose of this research paper is to review and comment on the psychology of these transient memories.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
Traditionally, memory researchers have distinguished between two types of transient memories: sensory memories and short-term memories. Sensory memories are faithful, veridical records of initiating stimuli. You can think of a sensory memory as a kind of continuation of the actual event—the same information simply removed in time (Crowder & Surprenant, 2000). By definition, then, sensory memories are modality specific: Visual stimuli lead to visual sensory memories, auditory stimuli lead to auditory sensory memories, tactile stimuli produce tactile sensory memories, and so on. Sensory memories tend to last for only a second or two, at best, and are widely thought to accrue from the processes involved in normal sensation and perception.
Short-term memories are the active, but analyzed, contents of mind. Any time that we form a conscious idea, or process incoming information from the world, we activate existing long-term memory structures; as a collective set, this activated knowledge defines what most psychologists currently mean by short-term memory (e.g., Cowan, 1995; Shiffrin, 1999). Short-term memories, unlike sensory memories, need not accurately reflect a just-presented stimulus. Instead, they usually represent meaningful interpretations of what has just occurred. For example, we might see a string of visual forms representing the letters P U M P K I N, but actively maintain a short-term memory for the word pumpkin and perhaps even a visual image of the orange object itself. Evidence suggests that short-term memories are often represented in the form of an acoustic code—an inner voice—which probably plays a vital role in the interpretation and production of spoken language (e.g., Baddeley, Gathercole, & Papagno, 1998).
Psychologists use an additional term, working memory, to refer to the set of processes, or systems, that control and maintain activation of short-term memories (Baddeley, 1986; Miyake & Shah, 1999). Activation is assumed to be inherently fragile, so short-term memories are quickly lost in the absence of some kind of maintenance process. The working memory system is thought to contribute to virtually all aspects of cognitive processing (e.g., reading, reasoning, problem solving, etc.), but this research paper focuses solely on the task of remembering over the short term. What are the characteristics of memory over the short term? How does memory over the short term differ from long-term memory? Are different systems or mnemonic principles needed to explain short- and long-term remembering? To begin, I turn to the briefest of memories: the lingering aftereffects of event presentation.
Prolonging the Present: Sensory Memory
It is easy to demonstrate that the internal experience of a briefly presented event outlasts the event itself. Twirl a sparkler on a warm summer night and you’ll see a trail of the light, perhaps enough to form a rough circle or to attempt the outlines of a name. The abrupt ending of a symphony, experienced in a quiet room, leaves an echo that contributes to the drama of the musical piece. The briefest of touches can linger, leaving behind a record of the preceding impression. These are sensory memories: fleeting, raw records of experience.
There is nothing in the concept of a sensory memory that is necessarily tied to the passage of time. We can recognize the sound of a person’s voice, or call to mind two viewings of the same visual scene, even though months or years might have passed. Most psychologists, however, use the term sensory memory to refer to stimulus persistence, a kind of prolonging of the present. Longer term modality-specific memories, such as the long-term recall or recognition of a person’s voice, are generally classified as perceptual memories to distinguish them from sensory persistence per se (e.g., Cowan, 1984, 1988; Massaro, 1975; Massaro & Loftus, 1996). There may be different forms of short-term sensory persistence. For example, some have suggested that there are two distinct phases of sensory storage, one lasting only a few hundred milliseconds and a second lasting up to 20 s (e.g., Coltheart, 1980; Cowan, 1984; Massaro, 1975).
Unfortunately, the distinction between the various forms of sensory persistence and perceptual memory is not a clean one and has led to some interpretive problems in the sensory memory literature. For example, many of the tasks that have traditionally been used to study sensory persistence may, in fact, be measuring perceptual memory (see Loftus & Irwin, 1998). Questions have also been raised about the adaptive value of the persistence process itself (Haber, 1983). In theory, a prolonging process seems clearly adaptive: To perceive a spoken word, it is necessary to integrate across phonemic information that is presented sequentially in time (Cowan, 1984; Darwin, 1976); integrating two visual images successfully, such as those produced by a rapidly moving object, may require one to maintain a relatively intact memory for the initially presented image (Eriksen & Collins, 1967). However, it is uncertain how much of a role sensory persistence actually plays in these situations, or whether it plays a role at all. Some researchers have suggested that sensory persistence may result from the fact that neural responses are simply extended in time (Loftus & Irwin, 1998; Francis, 1999). The fact that the subjective experience of persistence—even its very presence—depends on factors such as the duration and intensity of the physical stimulus supports this kind of explanation.
Measuring Sensory Persistence
As with most psychological phenomena, our understanding of sensory persistence has been largely defined by measurement techniques. One relatively direct technique, known as a synchrony judgment task, asks observers to adjust the timing of an index stimulus, such as an auditory click, until it coincides with the onset or offset of a target stimulus, such as a light (e.g., Bowen, Pola, & Matin, 1974; Efron, 1970). Observers are quite accurate at deciding when the target stimulus first appears—its onset—but overestimate its offset by around 150 ms; in other words, observers think the stimulus continues for a brief period after it has physically disappeared. Again, the extent of the persistence depends on the intensity and presentation duration of the target stimulus, but seems to be largely independent of presentation modality. For brief target presentations, both visual and auditory stimuli show sensory persistence effects lasting somewhere between 100 and 200 ms.
Comparable results are obtained using a technique called backward masking (e.g., Massaro, 1970; Turvey, 1973). Here, visual or auditory stimuli are presented and then followed closely by an interfering “mask” from the same presentation modality. The task is to identify or recognize the target stimulus. For example, in a simple auditory backward masking task a high- or low-pitched tone might be presented, followed at some variable time by another irrelevant but masking tone; the subject’s task is simply to categorize the pitch of the first tone (high or low). The common finding is that recognition or identification performance improves as the interval between the end of the target stimulus and presentation of the mask increases, until an asymptote is reached at around 250 ms (Massaro, 1970; see Figure 15.1). After 250 ms, further delays in presentation of the mask do not affect performance very much (see also Massaro & Loftus, 1996).
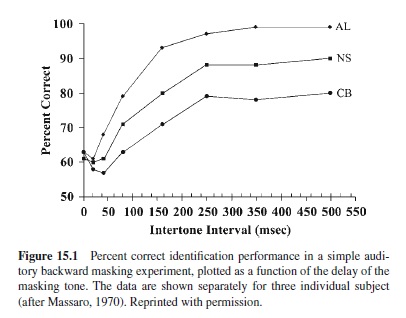
The 250-ms asymptote is commonly interpreted as the point beyond which the stimulus no longer persists—that is, the duration of the sensory memory. Alternatively, 250 ms could simply represent the point at which the subject has extracted all the relevant information that he or she needs. Many researchers find it compelling that similar masking functions are found for visual and auditory stimuli (Cowan, 1995; Massaro, 1975); furthermore, the estimated duration derived from backward masking—250 ms—roughly corresponds to the phenomenological duration tapped by the synchrony judgment task. Other persistence tasks yield similar duration estimates. For example, in a temporal integration task, subjects are asked to identify a missing element in an array of elements (e.g., Di Lollo, Hogben, & Dixon, 1994). Presentation of the array unfolds over time: Subjects receive a random half of the elements at Time 1 and the second half after a brief, but variable, delay. Successful performance requires perceptual integration of the two halves, which seems to occur only if the two halves are separated by fewer than 100–200 ms.
The Partial Report Technique
The measurement technique most commonly associated with sensory memory, particularly visual sensory memory, is the partial report procedure developed by Sperling (1960; see also Averbach & Coriell, 1961). When people are presented with a visual display of letters—for example, a three-by-four array of 12 letters—for a very brief duration—say, around 50 ms—roughly 3 or 4 letters can be reported correctly. People can presumably process only a limited amount of information in 50 ms, so this result may not seem surprising. However, people report the clear sensation of seeing the entire display, with all 12 letters, but the display fades before all the letters can be reported. Sperling (1960) set out to measure the persistence of the display, which he believed tapped a form of visual sensory memory.
Sperling’s first challenge was to document that people do, in fact, have more than three or four letters available following the offset of the display. He devised a partial report condition, which required the reporting of only part of the display rather than all 12 letters (hence the name partial report). After the display was turned off, one of three auditory cues sounded—a high-, medium-, or low-pitched tone— which signaled the subject to recall only one of the three rows of letters. Because the cue was presented after the display was physically terminated, average row performance could be used to estimate availability of the display as a whole. Sperling discovered that, indeed, people have much more display information available than the three to four letters tapped by whole report.
The partial over whole report advantage is important, but it does not, by itself, establish the presence of sensory memory. For one thing, fewer items need to be recalled under partial report (4 instead of 12 letters), so some kind of recall (or output) interference could be contributing to the condition differences. Of main interest is the finding that the partial report advantage declines rapidly with the insertion of a delay between display offset and the occurrence of the recall cue. Sperling found that the advantage was eliminated if the recall cue was delayed for 1 s after offset of the display, and it was sharply reduced after a few hundred milliseconds. The inference is that the visual display persists, as a sensory or iconic memory, for a brief period following offset, allowing the subject to continue processing its contents. Note that the estimate of duration derived from the partial report technique is slightly longer than, but in the same ballpark as, the estimates of persistence derived from the other procedures described previously.
Considerable work has been conducted using the partial report task over the past 40 years (see Greene, 1992; Massaro, 1975; Neath, 1998, for general reviews). Auditory versions of the task indicated initially that auditory sensory memory might last considerably longer than visual sensory memory (on the order of seconds rather than milliseconds; e.g., Darwin, Turvey, & Crowder, 1973), but methodological concerns cloud this conclusion (see Massaro, 1975). Other influential work has studied the particulars of the errors that occur in the task as the recall cue is delayed. It turns out that people primarily make location errors with delay: That is, the identities of the letter are not lost, but, rather, people become confused about the row the letters occupied (Mewhort, Campbell, Marchetti, & Campbell, 1981).
The Characteristics of Sensory Persistence
It is not yet certain whether each of the tasks just described really measures the same psychological construct—that is, a decaying sensory memory. Each has somewhat different task requirements, so it is reasonable to expect differences. For example, a synchrony judgment task does not require one to extract meaningful information from the display, such as letter identities, and the partial report technique requires one to allocate attention selectively to the cued location in the visual display (Dixon, Gordon, Leung, & Di Lollo, 1997). Few studies have attempted to compare the different techniques directly, but there is some indication that the partial report technique may be measuring something qualitatively different from what is tapped by the other techniques (Loftus & Irwin, 1998).
Inverse Effects
Most forms of sensory persistence, at least as studied by the majority of measurement techniques, do show common characteristics. For example, the duration of persistence is inversely related to target stimulus duration and intensity. Efron (1970) found that people were quite accurate at judging stimulus offset as long as the target stimulus was presented for at least 150 ms. In general, as the duration of the target stimulus increases, people experience less persistence. The duration of persistence also shortens as the intensity of the target stimulus increases: For example, brighter stimuli lead to shorter persistence effects. One interpretation of these results is that the target stimulus, at onset, initiates a period of neural activity that lasts for a few hundred milliseconds. If the physical stimulus is removed prior to the completion of this neural response, the stimulus will appear to persist until the response function is complete. If the duration of the stimulus itself exceeds the neural response time, or if some other factor, such as intensity, effectively shortens the neural response time, then no persistence effects will be found.
Neural Dynamics
At present, there is no consensus on the proper interpretation of these persistence effects. However, the idea that people have special sensory memory stores, designed to maintain literal copies of sensory input as an aid to subsequent perceptual processing, is losing favor among researchers. Instead, persistence effects are widely believed to result from the dynamics of neural processing, perhaps simply as an artifact of systems that are designed to accomplish more general ends. For example, Francis (1999) has proposed that visual persistence effects accrue from a general mechanism of excitatory feedback in cortical neural circuits; the duration of persistence, in turn, is driven by cortical “reset” signals that dampen, or inhibit, the feedback. Increases in stimulus duration or intensity increase the strength of the reset signals, thereby affecting the perceived duration of persistence (see also Grossberg, 1994).
To the extent that persistence effects are caused by general neural mechanisms, one might expect to find similar effects across all modalities. As discussed, there do indeed appear to be common performance characteristics across modalities, but more research needs to be conducted. Moreover, for methodological reasons, it is difficult to investigate persistence effects in some modalities; consider, for example, the inherent difficulties involved in controlling the presentation duration of an olfactory or gustatory stimulus. For this reason, little work has been conducted on modalities other than vision and audition. New techniques are in development, particularly techniques designed to tap neural processing (see Näätänen & Winkler, 1999), so answers may be on the horizon.
Modality and Suffix Effects
The evidence gleaned from the partial report technique, as well as the other measures of persistence, was used in the 1960s and 1970s to support the proposal of specialized sensory memory stores—that is, iconic memory for visual stimuli and echoic memory for auditory memory. These memory stores, in turn, were widely believed to contribute to performance in a number of perceptual and mnemonic tasks. In immediate serial recall, for example, a persisting auditory, or echoic, trace was thought to underlie the modality effect, which is the large recency advantage that one typically sees for lists presented aloud (Corballis, 1966). An extensive literature developed to explain how factors influenced the size of the modality effect, purportedly for the reason of understanding the characteristics of auditory sensory memory.
Precategorical Acoustic Storage
Crowder and Morton (1969) proposed that lingering echoic information was held in precategorical acoustic storage (PAS), a limited-capacity sensory store capable of holding a few auditory items. Unlike visual sensory memory, which decayed very rapidly, the contents of PAS were believed to last for several seconds, allowing a subject time to give the last one or two items in a memory list a kind of once-over prior to recall. This provided an end-of-the list advantage for aurally presented items because the subject could use the echoic information in PAS to correct selective information in short-term (or working) memory (see Crowder, 1976). A related empirical phenomenon, the suffix effect, provided supporting evidence: If an auditory list is followed by another redundant spoken item, such as the word recall, the auditory recency advantage is reduced or eliminated. In this case it was assumed that the suffix, because of the limited capacity of PAS, interfered with the echoic traces for the last list item or two, eliminating the recency boost.
Initially, a great deal of evidence accumulated supporting the PAS account. For example, delaying the suffix by a few seconds typically reduces its interfering effect, supporting the proposal that the contents of PAS have a useful lifetime of only a few seconds. In addition, the destructive power of the suffix depends importantly on its acoustic, rather than its semantic or categorical, similarity to the list items (J. Morton, Crowder, & Prussin, 1971). What is stored in PAS, according to the model, is a raw, uncategorized sensory trace; consequently, one would expect the modality and suffix effects to show sensitivity only to acoustic variables. Crowder (1978) further showed that if list items are comprised of homophones presented aloud (plus visually for identification), such as pare, pair, pear, no enhanced recency is found. In this case the lingering echoic information, although still stored in PAS, is nondiscriminative acoustically: It cannot be used selectively to correct the short-term memory records for recency items (see Nairne, 2001, for a fuller discussion).
In the 1980s, however, support for the PAS model was weakened considerably by the demonstration of modality effect patterns for nonacoustic presentation modes. For example, sharp recency effects were obtained for lip-read stimuli (Campbell & Dodd, 1980) and occurred when subjects silently mouthed visual stimuli (Greene & Crowder, 1984; Nairne & Walters, 1983). Neither lipreading nor mouthing involves sound, thus precluding a role for PAS, yet both produced serial position curves that mimicked those found for auditory presentation. The suffix effect was also discovered to be sensitive in some cases to conceptual attributes (Ayres, Jonides, Reitman, Egan, & Howard, 1979; Neath, Surprenant, & Crowder, 1993), and to last over intervals considerably longer than a few seconds (Watkins & Watkins, 1980). Although various attempts were made to rescue the PAS model from conflicting data (see Greene, 1992), the account generally has fallen into disfavor (see Neath, 1998). Instead, both the modality and suffix effects are now widely believed to be short-term memory phenomena, although some form of residual perceptual memory may play an important role (see Nairne, 1988, 1990).
Short-Term or Working Memory
Whereas sensory memories tend to be veridical copies of the environment, short-term memories comprise the stuff of immediate experience. Consider the process of remembering a telephone number as you cross the room. The numbers, no longer physically present, remain active in consciousness because you engage in a process of internal repetition (in what appears to be a kind of inner voice). If you are distracted prior to reaching the phone, or fail to rehearse, the numbers are likely to vanish, leaving you with considerable uncertainty— a number here or there perhaps, but little or no confidence about the final order.
This description, which corresponds to subjective experience, actually represents the standard way that most memory researchers think about remembering over the short term (see Nairne, 2002). Permanent knowledge structures are activated, creating short-term memories, which renders the activated information immediately and directly recallable. Because of inherent attentional and resource limitations, only a certain number of items can be refreshed, through rehearsal, prior to loss, creating the familiar limitations in memory span (e.g., Miller, 1956). The whole process is somewhat akin to a juggler’s attempt to maintain a set of plates in the air: The capacity of the juggler is determined by how well he or she can counteract the forces of gravity by effectively retossing each plate before it hits the ground (also see Nairne, 1996).
The standard model of short-term memory successfully explains a wide range of empirical data, everything from the recency effect in free recall to the intricacies of immediate serial recall (see Healy & McNamara, 1996, for a review). In recent years, various researchers have attempted to formalize the mechanics of how storage is controlled in the form of computer simulation models, and I review some of these models later in the paper. However, questions remain about the proper interpretation of how we remember over the short term. Not all researchers accept the standard juggler model of short-term memory, choosing instead to opt for general mnemonic principles that apply over both the short and the long term (e.g., Melton, 1963; Nairne, 2002). In the following sections, I review the empirical data base on shortterm retention with an eye toward shedding light on these controversies.
Forgetting Over the Short Term
Any discussion of short-term memory is properly begun with the topic of forgetting. As noted previously, it is the fact that we forget rapidly over the short term that produces the familiar limitations in memory span. The concept of a short-term memory capacity, in effect, is meaningful only because we typically fail to remember certain portions of a presented memory list. For many years, immediate retention was largely ignored by memory researchers. Instead, the focus was placed on multitrial learning, particularly paired-associate and serial learning, in the interest of specifying the conditions of transfer and interference (see Osgood, 1953). Single-trial immediate serial recall, which today reigns as the prototype of the short-term memory task, went largely unstudied for the first half of the twentieth century.
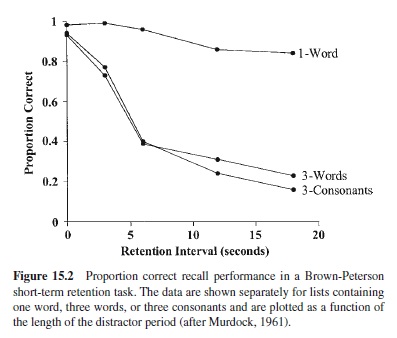
The situation changed dramatically around 1960 with the introduction of the Brown-Peterson technique, developed independently by John Brown (1958) and Lloyd and Margaret Peterson (Peterson & Peterson, 1959). In the original Peterson procedure, trials consisted of the presentation and recall of single consonant trigrams (e.g., CHJ) following a distractor-filled retention interval. Retention intervals varied randomly from 3 to 18 seconds and, importantly, the subject was required to count backwards aloud (by threes) throughout the interval to prevent rehearsal.The striking finding was that consonant trigrams, presented singly, were essentially forgotten after a retention interval of 18 s. Murdock (1961) extended the procedure to word recall and found similar results: nearly complete forgetting of a list of three words after approximately 18 s of counting (see Figure 15.2).
The Peterson finding was newsworthy for a number of reasons: First, most researchers were surprised to find any significant forgetting after such a short retention interval, especially given that the memory load was well below span. Second, the activity filling the retention interval—counting backward— lacked formal similarity to the to-be-remembered stimulus items (letters or words). Circa 1960, the main mechanism for forgetting was assumed to be interference, and not much interference was expected to occur between highly dissimilar materials. (There was some phonemic similarity between the letters and the digits, although this fact was not widely appreciated at the time.) Finally, the negatively accelerated form of the short-term forgetting curve showed a marked similarity to long-term forgetting functions (Ebbinghaus, 1885/1964), suggesting that it might be possible to study retention at a more fine-grained level (Slamecka, 1967).
Decay Versus Interference
Among the more interesting implications of this rapid shortterm forgetting, as noted by J. Brown (1958) and others (e.g., Broadbent, 1958), was the possibility that autonomous decay might be responsible for the loss. The notion that mnemonic information is lost spontaneously with the passage of time (e.g., as in Thorndike’s law of disuse) had largely fallen out of favor among psychologists, at least for long-term retention, because (a) memory sometimes improves with the passage of time (e.g., spontaneous recovery, reminiscence, or both) and (b) forgetting depends so critically on the nature of interfering material. John McGeoch’s famous analogy was of an iron bar left out to rust in open air: Rust accumulates with time, but it is the processes that operate in time (i.e., oxidation), not time per se, that are ultimately responsible for the changes (McGeoch, 1932). The fact that significant forgetting could occur in the absence of interference resurrected the concept of decay and bolstered the novel idea that short-term retention might be mediated by its own unique operating system.
As noted, retroactive interference could be easily dismissed as the source of short-term forgetting in the Brown-Peterson task, because counting and letters or words are highly dissimilar, but proponents of decay were forced to acknowledge that proactive interference might be responsible for at least some of the loss. Proactive interference is the interference that prior information, such as the items on trial N – 1, imposes on the retention of current trial information (trial N). Peterson and Peterson (1959) checked for proactive interference in their experiments but failed to find any support for it (in fact, performance actually improved from early to late in the session). However, in a landmark study, Keppel and Underwood (1962) eliminated practice trials and focused only on the very first trial. No proactive interference is possible on the first trial in a session, because there is no prior trial information, and Keppel and Underwood found almost no forgetting, regardless of the length of the retention interval. Differences between a short and a long retention interval began to emerge only on the second or third trial in the session, when, presumably, proactive interference was able to kick in.
Theoretically, the Keppel and Underwood (1962) findings are critical: If you believe in a separate short-term memory system, distinct from long-term memory, then it is important to show that short-term memory follows its own unique operating laws or solves problems that cannot be solved by the mechanisms governing long-term retention. The suggestion that most, if not all, short-term forgetting is caused by proactive interference—the same kind of interference that controls much of long-term retention—diminishes the rationale for rejecting a single-system view (see Melton, 1963). Other early work further supported the case for interference. Murdock (1961) found less forgetting when one word, rather than three, was used as the to-be-remembered stimulus (see Figure 15.2); in addition, Melton reported dramatically different short-term forgetting functions for lists varying from one to five consonants (a form of within-trial interference that he termed “intra-unit” interference). Others went on to show that even retroactive interference could play a role under some circumstances: For instance, more forgetting is found when items are presented aloud and the intervening distractor activity is also auditory (e.g., Proctor & Fagnani, 1978).
However, the fact that interference is operative in shortterm memory environments does not rule out decay; both decay and interference might be involved. Indeed, this was the position advocated a decade later by Baddeley and Scott (1971). They noted that in the Keppel and Underwood (1962) study, as well as in other studies documenting little effect of forgetting on the first trial (e.g., Cofer & Davidson, 1968), performance tended to hover near or at the ceiling. Thus, they argued, there might have been forgetting, but it was masked by the high performance levels. To get performance off the ceiling, they increased the length of the memory list from the standard three items to five items in one condition and seven in another. Under these conditions, significant forgetting was found on the first trial, but it appeared to reach asymptotic levels by around 5 or 6 s. Moreover, no differences were found in the slope of the forgetting curves as a function of list length, and the asymptotic levels of performance were significantly above the levels normally found when multiple trials are tested in a session. This suggested that some sort of decay operates early in the retention interval but is complete by around 5 s; it also suggested that interference, particularly from prior trials, must cause the bulk of the forgetting found later in the retention interval.
The conclusions reached by Baddeley and Scott (1971) have largely dominated the field for the past three decades. Most researchers believe that interference plays a significant role in short-term forgetting—in fact, interference is acknowledged to cause most, if not all, of the forgetting in the Brown-Peterson task—but few have completely rejected the concept of decay. As I discuss later, the case for decay was strengthened initially by the discovery of time-dependent limitations in short-term memory capacity (the word length effect; Baddeley, Thomson, & Buchanan, 1975). Moreover, it was subsequently shown that short-term forgetting is nearly complete after only a second or two if the recall test appears unexpectedly; in the traditional Brown-Peterson procedure, subjects expect the recall test and, consequently, may engage in elaborative processes that enhance long-term memory for the list items (see Healy & Cunningham, 1995; Muter, 1980; Sebrechts, Marsh, & Seamon, 1989). In the absence of elaborative processing, which enables a kind of back-up recall from long-term memory, one is forced to rely exclusively on the fragile activity trace, which decays rapidly—in a second or two—in the absence of rehearsal.
Temporal Distinctiveness
According to the standard model, as just discussed, decay of the activity trace is largely complete after only a few seconds. Interference, particularly proactive interference, is then largely responsible for any further forgetting that occurs during a retention interval. But what specific interference mechanisms are involved? One common assumption is that subjects are able to retrieve just-presented items from longterm memory, after the activity trace has decayed, but successful retrieval requires discriminating correct list targets from incorrect alternatives. Items from earlier trials, as well as extraexperimental items to a certain extent, form a noisy background against which the correct item must be selected.
There is a reasonable amount of evidence indicating that the mechanism for trace discrimination involves temporal or positional information (see G. D. A. Brown, McCormack, & Chater, 2001; Neath & Crowder, 1990). After all, time-ofoccurrence information, when available, provides a foolproof method for distinguishing items from Trial N from those occurring on the previous trial, N – 1. Various studies have shown that forgetting in short-term memory environments depends importantly on the temporal spacing of items within a list, the length of the retention interval, and the temporal spacing between trials in the session. It is not time per se that predicts retention, but rather the temporal relations among the items in the experimental session.
In a slight variation of the typical Brown-Peterson task, Turvey, Brick, and Osborn (1970) asked different groups of subjects to count backward as distractor activity for either 10, 15, or 20 s. Remarkably, no retention differences were found among these groups, despite the retention interval differences (see also Greene, 1996). Of main interest, though, was a critical trial in which all groups were switched to the same 15-s distractor interval. Correct recall dropped in the 10-s group (from .33 to .20), stayed roughly constant in the 15-s group (.30 to .28), and actually improved in the 20-s group (.30 to .38). Notice that the passage of time—and therefore the opportunity for decay—was equated across the groups on the critical 15-s trial, yet performance depended critically on the timing of prior trials.
According to distinctiveness accounts, memory improves to the extent that target items can be easily discriminated from the memories created by prior trials. As with most perceptual comparisons, the discrimination process is assumed to be relative: As time passes, temporally distant items seem more similar and become harder to discriminate, just as telephone poles watched from a moving car appear to merge together with increasing distance traveled (Crowder, 1976). Thus, two items separated by 5 s become harder to discriminate after 20 s than after 10 s. This relationship can be expressed easily in terms of a ratio: Discriminability is proportional to the interitem interval (the period separating the two items in question) divided by the retention interval (the time separating the most recent item from the point of test).
Now reconsider performance in the Turvey et al. (1970) experiment (see Figure 15.3). Prior to the 15-s critical trial, the intertrial intervals remained constant within the session (10, 15, or 20 s), creating a discriminability ratio of 1.0 for each group. As noted above, no retention differences were actually found among these groups, despite the different retention intervals, providing strong support for the distinctiveness account. On the critical trial, however, the ratios change differentially across the groups, in the direction predicted by the data. More specifically, the discriminability ratio declines in the 10-s group (10/15 = 0.67), leading to poorer retention performance, and increases in the 20-s condition (20/15 = 1.33), leading to improved performance; the ratio remains at 1.0 in the 15-s condition, and no performance changes were recorded (15 /15 = 1.0).

The Turvey et al. (1970) data are particularly important because they show that short-term memory performance can decrease, stay the same, or even increase depending on timing variables. Comparable results have been found in other contexts: Neath and Knoedler (1994), for example, found that memory for early items in a list sometimes improves as the length of the retention interval increases (see also Wright, Santiago, Sands, Kendrick, & Cook, 1985). According to distinctiveness accounts, early items become relatively more discriminable as all list items recede backward into the past (see also Bjork, 2001). No simple version of decay theory can handle such findings; the passage of time, it turns out, fails as a general predictor of short-term memory performance (except, perhaps, for the first few seconds of a retention interval).
Capacity Limitations
Obviously, to the extent that items are rapidly forgotten over the short term, for whatever reason, there will appear to be fundamental limitations in memory capacity. From the perspective of the standard juggler model outlined earlier, the storage capacity of short-term memory is determined by the trade-off between decay and the rate of internal rehearsal. The number of items that can be recalled correctly, in order, on at least half of the trials (i.e., memory span) is determined by the number of items that can be rehearsed within the timelimited decay window. That number, as noted by Ebbinghaus (1885/1964) and others (e.g., Miller, 1956), tends to be around seven plus or minus two unrelated items.
It is misleading, though, to think about capacity limitations in short-term memory simply in terms of items, particularly in terms of a number such as seven plus or minus two. First, as Miller (1956) showed, it is not really the number of nominal items but rather the number of functional “chunks” that influences span. We can remember seven unrelated letters, seven unrelated words, or even seven unrelated sentences with somewhat comparable degrees of efficiency (although see Cowan, 2001, for evidence that the limit may actually be closer to four). More importantly, though, a strong argument can be made that some other variable— perhaps time—truly controls retention. Holding the number of to-be-remembered chunks constant, immediate retention can vary dramatically as a function of item characteristics. For example, it turns out that memory span is well predicted by the length of time needed to say items aloud or repeat items aloud in succession.
The Limits of Time
In a seminal article, Baddeley et al. (1975) found that lists of short words lead to better immediate serial recall than lists of long words, even though the number of chunks (i.e., words) is constant across conditions. Of course, short and long words can differ in a number of ways (e.g., number of letters, syllables, and so on), so intraunit interference could conceivably account for this word length effect (e.g., Melton, 1963). However, Baddeley et al. (1975) found that the performance differences remained when all factors other than pronunciation time—or articulation rate—were controlled. For example, the words bishop and Friday have the same number of letters and syllables but differ in pronunciation time (Friday takes longer to say); lists of long words, defined solely in terms of duration, still yielded poorer recall than lists of matched short words.
As has been noted, the standard model assumes that decay is the main culprit behind forgetting, and the word length effect is certainly consistent with this view. The longer it takes to rehearse a set of items, the greater the chances that some of the items will be lost prior to refreshing. Pronunciation time, or articulation rate, is simply assumed to correlate with the speed of internal rehearsal. Specifying capacity in terms of items, then, is correct only in the sense that it usually takes more time to rehearse a large number of items. Baddeley et al. (1975) reported that the useful lifetime of a short-term activity trace is around 2 s (see also Schweickert & Boruf, 1986); consequently, memory span should be roughly equal to the amount of material that can be rehearsed in 2 s. On average, not surprisingly, we can rehearse somewhere around seven plus or minus two items in 2 s.
This relationship between pronunciation time and immediate memory span generally holds at the level of group data as well as for individual subjects. In fact, it is possible to predict individual differences in memory span, for both children and adults, by measuring overt articulation rate (see Hulme, Thomson, Muir, & Lawrence, 1984; Tehan & Lalor, 2000). Developmental changes in memory span are also associated with rehearsal rate, to a certain extent, as are some differences that occur in span cross-culturally. For instance, digit span tends to be higher in English and Chinese than in Arabic or Welsh, presumably because it takes longer to say digit names in the latter languages (see Ellis & Hennelly, 1980; Naveh-Benjamin & Ayres, 1986).
Recent data, however, are qualifying these conclusions somewhat. For example, it turns out that span differences sometimes remain even when pronunciation times are held constant. Memory span is typically lower for phonologically similar lists of words, compared to phonologically dissimilar lists, but similarity has little, if any, effect on pronunciation rate (Hulme & Tordoff, 1989; Schweickert, Guentert, & Hersberger, 1990). Hulme, Maughan, and Brown (1991) found that words can produce higher memory spans than nonwords, even when articulation rates are matched for the item types; similar dissociations between articulation rate and span have been found for concrete versus abstract words (Walker & Hulme, 1999) and for high- and low-frequency words (Hulme et al., 1997; Roodenrys, Hulme, Alban, & Ellis, 1994). Any model that appeals simply to time-based limits in storage capacity—for example, the standard rehearsal plus decay model—has no clear way of explaining these data.
Even more troubling are recent reports suggesting that one of the major conclusions of Baddeley et al. (1975)—namely, that duration-based span differences exist for word sets matched on all variables other than spoken duration—may apply only to restricted sets of words. Caplan, Rochon, and Waters (1992) found no memory advantage for shortduration words in lists containing short- and long-duration words matched for number of syllables and phonemes; instead, they reported a reverse word length effect (long better than short) when duration was implemented by using “lax” vowels of short duration (e.g., carrot) and “tense” vowels of long duration (e.g., spider). Caplan et al. (1992) suggested that the phonological structure of a word, not its spoken duration, determines the magnitude of the word length effect. A similar conclusion was reached by Service (1998) using Finnish stimuli, which allow one to vary duration by manipulating combinations of the same articulatory and acoustic features. Lastly, Lovatt, Avons, and Masterson (2000) varied spoken duration in disyllabic words, holding constant a host of potentially confounding factors (e.g., frequency, familiarity, number of phonemes) and found no advantage for shortduration words across several experiments; word duration effects emerged only when the original word sets used by Baddeley et al. (1975) were used as the to-be-remembered stimuli.
The Limits of Attention
In retrospect, it is not surprising that factors other than time contribute to limitations in immediate retention. Even within the standard model, storage capacity is not fixed, but rather arises from the trade-off between decay—which is purely time-based—and a controlled process of rehearsal. Successful retention depends on one’s ability to keep list items in an active and recallable state, but also on the ability to counteract distraction and eliminate interference from nontarget information in memory. Errors in immediate retention, for example, often turn out to be intrusions from immediately preceding trials (e.g., Estes, 1991; Wickelgren, 1967).
Some researchers believe that limits in immediate memory arise, at least in part, from the ability to use controlled attention to ignore or filter out potentially interfering material (see Dempster, 1992; Kane & Engle, 2000). There is some evidence to suggest that individuals with low memory spans are more susceptible to proactive interference than high-span subjects: Rosen and Engle (1998), for instance, found that low-span subjects were more likely to intrude previously learned items into a current paired-associate recall task. It is also possible to get high-span subjects, who presumably possess more capacity for controlled attention, to mimic lowspan subjects’ susceptibility to interference by having them perform an additional concurrent task (Kane & Engle, 2000). The greater the amount of controllable attention, the easier it is to inhibit or reject interfering material as well as to keep target items active in memory through rehearsal.
It is also worth noting that some measures of capacity correlate reasonably well with other cognitive measures, such as reading comprehension, vocabulary learning, and even intelligence. For example, Engle and colleagues developed the operation span task, in which the presentation of to-beremembered items is interspersed with a requirement to solve simple addition problems (e.g., Turner & Engle, 1989). The number of words recalled is still of main interest, but the dual task conditions (arithmetic plus immediate retention) seem to tap attentional capacity to a greater extent than simple span measures. The operation span task, as well as related measures (e.g., Daneman & Carpenter, 1980), turns out to predict higher order cognitive abilities such as general fluid intelligence or the verbal scholastic aptitude score (see Engle, Kane, & Tuholski, 1999, for a review).
What emerges is a view proposing that the storage capacity of short-term memory, as defined generally by a measure such as memory span, is determined by a variety of factors, not a single factor such as a magic number of seven plus or minus two. The capacity to focus and sustain attention, engage in strategic rehearsal, and even recall quickly (Dosher & Ma, 1998) modulates the number of items that can be remembered over the short term. The characteristics of the items also matter: Word frequency, imageability, and lexical status all influence memory span, as does the similarity among the items presented together in a list. Another important factor is the rhythm and timing of stimulus presentation: If temporal gaps occur predictably within list presentation, immediate memory can improve substantially (e.g., Hitch, Burgess, Towse, & Culpin, 1996; Ryan, 1969). All of these factors need to be explained by a complete model of immediate retention.
It is also the case that any act of remembering will be influenced by the nature of the retrieval environment, regardless of whether the remembering occurs over the short or long term. As I discuss in the next section, retrieval from short-term memory, like long-term memory, is essentially cue-driven. Moreover, the effectiveness of cues depends on how target information has been encoded, as well as the extent to which the cue uniquely specifies the to-be-remembered item. This means that even with unlimited amounts of time, or an unlimited amount of attentional capacity, there can still be forgetting and, therefore, apparent limitations in storage capacity. Although there may be a relatively fixed amount of resource or attentional capacity available at any moment in time, understanding this limit will not explain, or effectively predict, all instances of short-term retention.
Retrieval of Short-Term Memories
As defined earlier, short-term memories are the active, but analyzed, contents of mind. By virtue of their activation, some researchers have assumed that they are immediately available for recall—that is, short-term memories exist in a state that allows for direct and effortless retrieval (e.g., McElree, 1998; Wickens, Moody, & Dow, 1981). On reflection, however, it is difficult to see how such a mechanism for remembering might actually work. For one thing, multiple short-term memories exist concurrently (short-term memory, as a whole, is often described as the set of activated knowledge), so a mechanism is needed to select a particular activated item for recall. More importantly, as just noted, the success of immediate retention seems to depend critically on the nature and extent of retrieval cues that are available.
Most recent models of short-term retention assume that the short-term activity trace forms the basis of immediate memory, but the trace needs to be interpreted, or deblurred, prior to actual recall. Interference, or possibly decay, degrades the activity trace over time, rendering its identity equivocal. The term redintegration is widely used to describe the interpretation process, which is assumed to rely on information stored in long-term memory. It is here, during the redintegration stage, that item characteristics such as word frequency or concreteness probably exert their effects. For example, one can assume that time-dependent rehearsal affects the intactness of the activity trace at the point of recall, but item-based characteristics (e.g., concreteness) affect the ease of redintegration (see Schweickert, 1993). Separating the status of the activity trace itself from its interpretation prior to recall allows one to explain, for instance, how immediate recall differences can occur despite the equating of pronunciation time.
Retrieval Dynamics
Assuming that a set of activated information exists at any moment in time, how does one select an appropriate candidate to recall? In the 1960s, Saul Sternberg developed a task to investigate the retrieval process. In Sternberg’s task, subjects are presented with short, below-span lists of items (e.g., words, letters, digits) followed immediately by the presentation of a single probe; this probe either matches, or not, one of the just-presented items (e.g., Sternberg, 1966). Obviously, a task of this kind is relatively easy, and people rarely make mistakes. Of main interest is the latency, or reaction time, of correct responses, usually as a function of a variable like list length.
Sternberg found that mean reaction time increased linearly with list length, but the slopes of the reaction time functions were roughly equivalent for positive and negative responses. This suggested that people search short-term memory in a serial, or item-by-item, fashion looking for a match to the recognition probe; the more items that need to be searched, the longer the reaction times. Equal positive and negative slopes suggested as well that the search process was exhaustive, meaning that all the items in the set were compared regardless of when (or whether) a match was found. The proposed serial exhaustive search process seemed to rule out other plausible search procedures—for example, selfterminating (in which the search process stops once a match is found) or parallel processing (in which all activated items are compared simultaneously with the probe).
However, it turns out that mean reaction time cannot be used to discriminate definitively between serial and parallel search processes; it is possible to mimic the reaction time patterns noted by Sternberg (1966), for example, using a parallel processing mechanism with certain additional assumptions (e.g., Townsend & Ashby, 1983). More diagnostic evidence comes from either a fuller analysis of reaction time distributions (e.g., Ratcliff, 1978; Reed, 1976) or from techniques that examine the full time course of processing during recognition decisions. In the latter instance, the response-signal procedure cues the subject to respond at particular times after the appearance of the recognition probe: For example, the cue to respond might appear almost immediately after appearance of the probe, which yields performance near chance levels, or seconds later, which allows for the most accurate performance. One can then determine how accuracy unfolds over time—so-called retrieval dynamics—which allows for a more sensitive analysis of possible retrieval mechanisms (see Dosher & McElree, 1992; McElree & Dosher, 1989).
Application of the response-signal technique to the retrieval of short-term memories in the Sternberg task supports a parallel, direct-access retrieval process. Retrieval dynamics seem not to vary much with list length, or serial position, which is consistent with a parallel matching process (Ratcliff, 1978). The nature of the retrieval process may change, however, depending on the type of information that must be retrieved. McElree and Dosher (1993) report that the recovery of order information—which of two list items occurred more recently?—is accomplished through a slow serial retrieval process; again, this conclusion is based on the finding that the retrieval dynamics for the order judgment differ systematically from those found in item recognition (i.e., the Sternberg task). The fact that the retrieval dynamics vary in this way undermines the simple notion that activity traces exist in a state of immediate availability. Even if items exist in a special focus of attention (e.g., Cowan, 1995), by virtue of their activation, various kinds of retrieval-based selection processes are clearly needed to satisfy the demands of differing tasks.
It is possible, however, that there is something special about retrieval of the very last item, or item chunk, in a short list. The last item is recognized faster than other items, but more importantly, the retrieval dynamics appear different as well (McElree & Dosher, 1989; Wickelgren, Corbett, & Dosher, 1980). This finding has been interpreted to mean that the last item remains active in awareness and, thus, can be matched directly with the recognition probe; the item essentially remains in consciousness, eliminating the need for a retrieval mechanism to move it from a passive to an active state. McElree (1998) recently showed that up to three items at the end of the list can show these special properties, as long as they are members of the same category (forming, presumably, a category chunk). However, alternative interpretations of the data pattern are possible. For example, one could argue that the contextual cues available at the point of probe presentation especially match the cues associated with the last list item; this, in turn, could affect the ease and quality of the retrieval process.
Cue-Driven Retention
One of the troubling features of direct access (or cueless retrieval) is the idea that items can be remembered without considering the nature of the retrieval environment. As Tulving (1983) has argued, there is no justification for making absolute statements about the memorability of items—for example, based on their inherent characteristics or encoding properties—because remembering always depends on an interaction between encoding conditions and retrieval conditions. It is possible that short-term memories represent a special case, violating Tulving’s dictum, but the available evidence suggests otherwise.
Direct access received some early support from studies showing that immediate retention, tested without a distractor interval, can show immunity to proactive interference (Halford, Mayberry, & Bain, 1988; Tehan & Humphreys, 1995; Wickens et al., 1981). In the Tehan and Humphreys (1995) experiments, trials consisted of the presentation of either one or two short four-item lists; the subject’s task was to recall the last presented list, so on two-list trials subjects were told to ignore the first list. Of main interest was the effect of the first list on recall of the second list—that is, proactive interference. On interference trials, one or more items in the first list were drawn from the same taxonomic category as items in the second list; on control trials the items were unrelated across the lists. When subjects were tested immediately, Tehan and Humphreys found no evidence for proactive interference (recall of control trials was equal to interference trials), but a significant control advantage emerged when a 2-s distractor task (reading digits aloud) occurred prior to recall.
If short-term memories are immediately available in consciousness, requiring no cue-based retrieval, then there is no reason to expect interference from a prior trial: One does not need to discriminate current items from previous items because the former are still active in awareness. However, immunity from interference turns out to be illusory. Semantic similarity between the first and second lists fails to hurt performance only because, on immediate tests, subjects can use residual phonemic information from the second list as a cue to help discriminate second- from first-list items. If the first and second lists contain items that rhyme, immunity disappears and significant proactive interference is found on the immediate test (see Tehan & Humphreys, 1995, 1996, 1998). Presumably, these subjects still have residual phonemic information available at test, but that information no longer uniquely specifies items from the second list.
Existing evidence strongly supports the idea that immediate retention is cue-driven. First, as described earlier, immediate recall is very sensitive to item characteristics, such as word frequency, lexicality, and concreteness. Most researchers assume, as a result, that the short-term activity trace must be interpreted prior to recall—that is, the short-term activity trace acts essentially as a cue to guide retrieval. Second, there is a considerable literature on release from proactive interference that confirms the importance of cues in immediate retention. In a typical release experiment, successive lists are drawn from the same conceptual class (e.g.,fruit). Recall gets worse over trials, presumably because people have a difficult time discriminating items on the current trial from conceptually similar items that occurred on previous trials. On the release trial, however, list items are drawn from a new conceptual class (e.g., moving from fruit to animals) and performance improves substantially (see Wickens, 1970).
This effect is most commonly interpreted to mean that people are using conceptual class as a cue to guide short-term recall; the effectiveness of this cue, in turn, hinges on its ability to predict current trial information. When successive lists are presented from the same conceptual class, the cue becomes overloaded (Watkins & Watkins, 1975) which means it starts to predict many items, especially those from previous trials. On release trials, however, the distinctive power of the cue is regained: It now uniquely specifies information from the current trial, and performance improves. Particularly strong support for this interpretation comes from experiments in which the nature of the retrieval cue is manipulated at test, after the critical list has actually been presented. It is possible to record significant levels of release, at test, if discriminating cues are provided (see Dillon & Bittner, 1975; Gardiner, Craik, & Birtwistle, 1972).
Diagnostic evidence for cue-driven retention also comes from the study of errors in immediate recall. When mistakes are made, the errors that occur tend not to be random, but rather follow certain patterns. For example, when a list item is recalled in an incorrect serial position, it is typically placed in a nearby position (e.g., the third item on the list is placed incorrectly in the second or fourth serial position). Recorded error gradients are systematic, showing that incorrect item placements drop off regularly as distance from the original position of occurrence increases (e.g., Healy, 1974). If lists are grouped, and people wrongly place an item from one group into another group, the item tends to be put in an identical relative serial position (e.g., Henson, 1999). Finally, when people intrude an item from a previous list, it is likely to have occurred at the same serial position in that list (Estes, 1991; Henson, 1996). Data of this sort indicate that people are not simply outputting activated items from short-term memory, but rather may be using some kind of position cue to help decide what occurred on the just-presented list.
Clearly, any full understanding of short-term memory will require some specification of how short-term memories are retrieved and translated into performance. The idea that immediate retention is cue-driven is appealing, in particular, because it is consistent with how most researchers conceive of long-term retention. All forms of remembering are cue-driven, although the nature of the cues, as well as the mechanics of the retrieval process, may differ between the short and the long term. This means that any empirical dissociations between short- and long-term retention, by themselves, will not be diagnostic of separate short- and long-term memory systems. One could still hold to a single system view, in which all forms of remembering are guided by the same set of general operating principles, and simply attribute the dissociations to differences in the composition of active retrieval cues (see Nairne, 2002).
The Working Memory Model
In the remainder of the research paper, I discuss some popular theoretical conceptions of the short-term memory system, beginning with the working memory model championed by Alan Baddeley (e.g., Baddeley, 2000; Baddeley & Logie, 1999). The working memory model was developed initially to counter the view that short-term memory is a unitary storage system: a single place, or store, where complex forms of cognitive processing (e.g., reasoning or language comprehension) occur concurrently with temporary storage. Baddeley and Hitch (1974) argued instead for a multicomponent system with separate subsystems designed to handle particular kinds of processing, such as the temporary storage of visual versus phonological information. The working memory model maintains a strong distinction between short- and long-term memory, but it fractionates short-term memory into separate parts.
Baddeley and Hitch (1974) noted, for example, that remembering a span-length list of items produces little disruption of a concurrent reasoning or problem-solving task. If both temporary storage and on-line cognitive processing are controlled by the same processing machinery—the same processing store—then significant interference should have occurred between the two. The fact that little interference is found suggests that temporary storage and attention-based central processing may be controlled by separate mechanisms. Data from the study of brain-damaged patients proved troubling as well: It was discovered, for example, that patients with severely impaired short-term memory can show relatively intact long-term memory (e.g., Shallice & Warrington, 1970); a view proposing that both temporary storage and long-term learning are controlled by the same system has trouble accounting for this pattern.
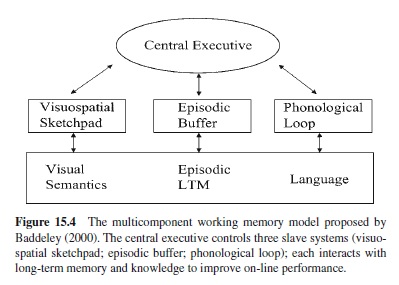
The working memory model has undergone significant changes since its inception, but its core architecture still consists of three basic components: the central executive, the phonological loop, and the visuo-spatial sketchpad. The central executive, as the name suggests, controls and coordinates the actions of the remaining subsystems. It is assumed to be a limited-capacity attentional system that directs the focusing and switching of attention, and it may play a role in activating structures in long-term memory as well (see Baddeley, 1996). The central executive plays no role in storage per se, except as the controller of the loop and the sketchpad. The central executive is the least well-specified working memory component and, as Baddeley readily admits, it often serves as a kind of theoretical “grab bag” for intractable problems (see Baddeley, 2000).
The Phonological Loop
The bulk of the empirical effort on working memory has been spent on the phonological loop, which is the system assumed to control the temporary storage of acoustic and verbal information. The loop is divided into two components: a phonological store, which is the actual storage location for to-be-used information, and a rehearsal/recoding device called the articulatory control process. Information residing in the phonological store decays in roughly 2 s, although it can be refreshed, via rehearsal, through the articulatory control process. Capacity limitations in immediate retention— for example, the magic number seven—are assumed to arise from trade-offs between decay and loop-based rehearsal. In essence, the phonological loop account is a prototypical instantiation of the standard juggler model described earlier in the paper (see Nairne, 1996, 2002).
The success of the working memory account hinges on its ability to explain a wide range of standard empirical phenomena. For example, the loop provides a nice account of the word length effect, discussed earlier, by assuming that there are inherent limitations in the operation of the articulatory control process. Memory span is limited to roughly what a person can rehearse within the time window established by decay—that is, about 2 s. When words are long, fewer can be refreshed before decay renders the short-term memory traces unreadable. The model also successfully predicts that the word length effect should be eliminated under conditions of articulatory suppression (Baddeley et al., 1975). Articulatory suppression—repeating a redundant item (e.g., the) aloud— acts to block rehearsal, thereby eliminating the mechanism that produces the word length effect.
In addition to refreshing decaying activity traces, the articulatory rehearsal mechanism serves an additional recoding function: It translates verbal material into phonological form. Representingstoredtracesphonologically,inthephonological store, enables the model to handle the phonological similarity effect, the finding that lists containing similar-sounding items (e.g., g, c, b, t) are harder to recall than lists of dissimilarsounding items (Conrad, 1964). Moreover, by linking the phonological translation process to rehearsal, the model generates the unique prediction that the phonological similarity effect should be eliminated under articulatory suppression, at least for visually presented material. Preventing rehearsal blocks the recoding function, forcing one to rely on nonphonological forms of storage. In fact, articulatory suppression does seem to eliminate the phonological similarity effect when materials are presented visually (e.g., Murray, 1968). When list items are presented aloud, the effect remains under suppression, presumably because auditory materials are automatically registered in the phonological store (see Baddeley, 1986).
Along with its temporary storage functions, the phonological loop is also assumed to play a very important role in language processing, particularly the learning of new phonological material. Variables known to affect the functioning of the phonological loop, such as articulatory suppression and the word length effect, also affect one’s ability to learn novel phonological forms, such as those required in the learning of a second language (see Baddeley et al., 1998). The learning of new words by children can also be predicted reasonably well by nonword repetition, a task that is assumed to tap functioning of the phonological loop. Finally, patients who show severe impairments in short-term memory tasks but show generally intact long-term memory and learning appear to have a selective deficit in the long-term learning of phonological information (Papagno, Valentine, & Baddeley, 1991). Baddeley et al. (1998) argue that the phonological loop may have evolved primarily to store unfamiliar sound patterns during time periods when more permanent memory records are being constructed.
The Visuo-Spatial Sketchpad
Whereas the phonological loop handles the temporary storage of verbal and acoustic information, the visuo-spatial sketchpad controls short-term processing and retention of visuospatial material. Like the phonological loop, the sketchpad probably has separable components, controlling visual, spatial, and possibly kinesthetic information (Baddeley, 2000; Baddeley & Logie, 1999). The visual component, which helps to retain visual patterns, is known as the visual cache; the capacity to remember sequences of spatial movements is attributed to an inner scribe (see Logie, 1995).
Most of the research investigating the sketchpad has employed dual-task methodologies. The goal is to demonstrate selective interference, thereby dissociating the capacity to retain visual, spatial, or verbal information, or combinations of these types of information. If subjects are asked to learn a list of words using an imagery mnemonic, which presumably taps the sketchpad more than the phonological loop, performance is hurt by the concurrent requirement to track a moving spot of light; the same tracking task has little, if any, effect on performance when subjects use a verbal-based rote learning strategy (Baddeley & Lieberman, 1980). Changing, but irrelevant, visual materials have been shown to disrupt the short-term retention of visual information (e.g., Quinn & McConnell, 1996); the retention of spatial patterns can also be selectively disrupted by spatial movements during a retention interval (see Baddeley & Logie, 1999). Collectively, these dissociations bolster the case for proposing separate storage mechanisms for verbal, spatial, and visual information.
However, at this point, there is no firm consensus on the inner workings of the sketchpad. It is unclear how dissociable the visual cache and the inner scribe will turn out to be, or the extent to which the different components draw on the same cognitive resources. Questions have also been raised about the relationship between mental imagery, in general, and operation of the sketchpad. Some evidence suggests that the two are dissociable. For instance, patients have been discovered who perform poorly on mental imagery tasks (such as mental rotation) but handle the short-term retention of visuo-spatial information quite well (N. Morton & Morris, 1995). In addition, some concurrent tasks, such as arm movements, that selectively disrupt the retention of spatial patterns have little effect on the performance of mental imagery tasks (Baddeley & Logie, 1999).
The Episodic Buffer
The working memory model has been enormously influential as an explanatory heuristic. It successfully ties together a wide range of standard laboratory phenomena, as well as data gathered from developmental and neurological studies. The model does have inherent problems, however, which it shares with other implementations of the standard juggler model (see Nairne, 2002, for a full discussion). For example, as noted earlier, word duration is probably not the important controlling factor in the word length effect. Words matched for pronunciation duration, but differing along other dimensions such as lexicality, regularly lead to memory span differences. The working memory model has no obvious mechanism to handle such effects. Moreover, the phonological similarity effect is assumed to result from confusions among representations in the phonological store, but no mechanism has ever been offered to explain exactly how these confusions arise. If items are immediately available by virtue of their residence in the store, why do the confusions occur? Is there some cue-based retrieval mechanism in place that can explain phonological confusions as well as other cue-driven immediate retention effects?
Another issue that has particularly troubled Baddeley is the question of how verbal information is stored temporarily when the phonological loop is unavailable. For example, under articulatory suppression, immediate memory performance is impaired, but only slightly: That is, performance might drop from a span of seven items to a span of five (Baddeley, Lewis, & Vallar, 1984). Given that the phonological loop is filled to capacity by the suppression activity, how are these items being stored? One possibility is the central executive, but Baddeley has assumed that the central executive performs no storage function (see Baddeley, 2000). Another possibility is the sketchpad, but verbal materials show little sensitivity to visual similarity under articulatory suppression. As Baddeley has recently stated, “the data suggest the need for some kind of ‘back-up’ store that is capable of supporting serial recall, and presumably of integrating phonological, visual, and possibly other types of information” (Baddeley, 2000, p. 419).
To solve these and other problems, Baddeley recently proposed a new working memory component—the episodic buffer—to serve as a limited capacity temporary storage system (see Figure 15.4). Controlled by the central executive, the episodic buffer differs from the loop and the sketchpad in performing both a storage and an integrative function; information from many different sources can be tied together in the buffer, including semantic information, and the result is a multidimensional episodic code. Presumably, the buffer enables one to store material when the loop or the sketchpad is unavailable, and it helps to explain how certain item characteristics (e.g., lexicality or imageability) might affect remembering over the short term. It is difficult to judge the merits of this new component of working memory at this point, however, because it has little, if any, unique empirical support.
Simulation Models of Short-Term Memory
Over the years, a number of formal simulation models (either mathematical or computer-based) have been proposed to explain the particulars of short-term retention. Probably the best known is the Atkinson and Shiffrin (1968) buffer model, which maintained a distinction between the structural features of a memory system (e.g., a limited-capacity short-term store) and the strategic control processes that operate within those structures (e.g., rehearsal, coding, or both). The buffer model established the mold for many subsequent modeling attempts, but most current models possess a decidedly different flavor. For example, whereas little attention was given in the Atkinson and Shiffrin model to the retrieval of short-term memories (items were simply dumped out of the short-term store), most current efforts focus extensively on the retrieval and interpretation of activity traces.
Virtually all current models maintain the distinction between short-and long-term memory,but they differ in whether similar processes are assumed to operate in the two cases. For example, some models essentially mimic the standard juggler model and attribute many of the standard immediate memory phenomena to a trade-off between rehearsal and decay (e.g., Burgess & Hitch, 1999; Henson, 1998; Page & Norris, 1998). Other so-called unitary models reject the concept of decay and offer little role for rehearsal, assuming instead that short-term retention is controlled by the same processes that control all forms of remembering—that is, both short- and long-term (e.g., Anderson & Matessa, 1997; G.D.A.Brown,Preece,&Hulme,2000;Nairne,1990).Space does not permit a full accounting of these models, but I very briefly outline some of their main features in the following two sections.
Hybrid Models
I use the term hybrid to classify the first set of models, because most acknowledge the important role that cue-driven retrieval processes play in short-term retention. Thus, items are not dumped out of a short-term buffer or loop; instead, recall candidates are chosen through some kind of item selection mechanism. This is a characteristic shared by unitary models that assign no special properties to remembering over the short term. In most other respects, though, hybrid models are simply implementations of the standard juggler model: Performance is based on short-term activity traces that are subject to immediate decay in the absence of continued internal rehearsal.
In the primacy model of Page and Norris (1998), immediate retention of serial order is controlled by the relative activation levels of list item traces. Activation level is determined by a primacy gradient, such that the trace for the first list item is assumed to be more active than the second list item, and so on. At the point of recall, items are selected for output based on their activation level, which means that the first list item tends to be output first and then suppressed. The output selection process is noisy, so there is a certain probability that items will be selected for output out of their proper sequence (leading to errors). Page and Norris (1998) have shown how these simple assumptions, along with standard assumptions about the trade-off between decay and rehearsal, can produce serial position functions and error gradients that mimic the patterns normally found in short-term serial recall.
In many respects, the primacy model attempts to formalize the main features of Baddeley’s phonological loop. By adding specificmechanisms—forexample,primacy-basedactivation gradients,noisyselectionprocesses,suppression—itbecomes possible to generate precise and sometimes novel predictions. For example, the primacy model predicts a unique and interestingkind of output error calledafill-in error. Itturnsout that when people miss an item in the recall sequence, as when they fail to recall the letter B in the second serial position, there is a significant tendency to report the missed item in the next serial position (i.e., as occurring in Position 3). Page and Norris (1998) argue that the assumptions of the primacy model correctly predict this tendency, whereas other models do not (although see Neath, Kelley, & Surprenant, 2001).
The connectionist model of Burgess and Hitch (1999) also attempts to formalize the operation of the phonological loop. Serial recall is more clearly cue-driven in this model, rather than based on activation levels, and the cues in this case are elements of a moving or drifting context signal. List items are associated with a snapshot of the context, which is set at the moment an item is presented. Adjacent items, because the context signal is slow-moving, tend to be associated to similar contextual cues, which helps to explain, in part, why people tend to transpose adjacent items in recall output. At the point of recall, the context signal is reset, and a competitive selection process proceeds based partly on the activations that items receive via their connections with the context.
The Burgess and Hitch (1999) model also assumes that connections exist between phonemic information and to-berecalled items. This phonemic layer, when activated, serves essentially as the analogue for the phonological store in the working memory model. Rehearsal strengthens otherwise decaying connections between the phonemic layer and items, thereby helping to account for phenomena such as the word length effect. Again, the details are beyond the scope of this research paper, but the model nicely handles a variety of phenomena in immediate retention, including the occurrence of soundbased errors, the effects of articulatory suppression, and even temporal grouping effects (see also Hitch et al., 1996).
Another recently developed hybrid model is the Start-End model of Henson (1998). Once again, the major assumptions of the standard juggler model are reproduced in the form of decaying representations that are refreshed through internal rehearsal. The locus of the word length effect, and the general relationship between articulation rate and span, is placed in the trade-off between rehearsal and decay. The unique aspect of the Start-End model is its machinery for handling the recovery of serial order. Henson (1998) assumes that, during presentation, list items are coded relative to the start and end of the list. Items near the beginning of the list are associated more strongly with a beginning-of-the-list start marker, and recency items more strongly with an end marker. These position codes are then reinstated at test and used to activate associated items, and an item is selected for recall.
Because of the nature of the position codes, adjacent items tend to be associated with overlapping cues, leading to systematic error gradients in recall. The Henson (1998) model, like the other hybrid models, can be shown to mimic the major phenomena of immediate retention.
Unitary Models
The second class of simulation model, the unitary models, typically rejects decay and rehearsal as the major determinants of immediate memory performance. As noted earlier in this research paper, there are both empirical and theoretical reasons to question whether decay and rehearsal are viable explanatory constructs in immediate retention. Although people certainly do rehearse, and rehearsal can play a role in unitary models, its role in unitary models is not to refresh otherwise decaying representations. Instead, rehearsal is typically viewed as another kind of stimulus presentation, which can, depending on the circumstance, either facilitate or interfere with subsequent retention (see G. D. A. Brown et al., 2000; Tan & Ward, 2000).
In the OSCAR model (which stands for OSCillator-based Associative Recall), forgetting is caused entirely by various forms of interference (G. D. A. Brown et al., 2000). By relying on interference and rejecting decay, OSCAR shares an important property with most conceptions of how forgetting occurs in long-term memory (see Crowder, 1976; Neath, 1998). In a fashion similar to the Burgess and Hitch (1999) model, associations are formed between to-be-recalled items and snapshots of a moving context signal (instantiated through sets of slow and fast temporal oscillators). The context is reset for recall, and there is cue-driven competition for output. Interference in the model occurs because of response competition during the selection process, output interference, and inherent capacity limitations in the storage mechanism that is employed (see Brown et al. for details).
Regarding rehearsal, the OSCAR model includes no active mechanism for the rehearsal of items during list presentation. Although its authors did consider the possibility of strategic rehearsal, the concept was rejected because, essentially, “we have found no need for it in accounting for the phenomena under consideration” (G. D. A. Brown et al., 2000, p. 172). With regard to decay, the concept is rejected for many of the same reasons discussed earlier in this research paper—that is, certain data seem antagonistic to the proposal of decay—but also because OSCAR is designed to explain data from both short-and long-term retention environments. Indeed, OSCAR fits data from retention intervals lasting seconds as well as it fits data from intervals lasting hours, without changing any of its main assumptions.
A similar characteristic is found in the perturbation model of Estes (1972, 1997), an early simulation model that strongly influenced the development of OSCAR. In the perturbation model, items are effectively represented as values along dimensions, such as temporal or spatial position. The organization can be hierarchical, meaning that an item might be represented in terms of its position along an ordered list, within-list, or within-group dimension (see Lee & Estes, 1981; Nairne, 1991). The crux of the model is the assumption that the position values are subject to random perturbations over time: That is, there is a certain probability that each represented value will drift along its position dimension. The probability that a perturbation will occur can be specified mathematically, and the model has been shown to generate precise predictions about correct and incorrect performance.
For example, the model does an excellent job of explaining the nature of errors in immediate serial recall. As noted earlier, when people make errors in ordered recall, items tend to be placed incorrectly in nearby positions, and there is a regular gradient found as distance increases from the item’s original position (see Healy, 1974). The perturbation model not only generates these errors, but it also specifies exactly how the gradients should change as retention intervals increase. The model also nicely handles empirical dissociations between item and order memory, particularly the different forms of the serial position curve that have been reported (see Healy, 1974). Moreover, as Nairne (1991, 1992) has shown, essentially the same assumptions that handle data from immediate serial recall can also fit data across retention intervals lasting minutes or hours (although see Healy & McNamara, 1996, for some qualifying arguments). Thus, the perturbation model can be viewed as a unitary model, explaining both short- and long-term memory performance, and neither rehearsal nor fixed decay assumptions are needed to fit the data (Estes, 1997; Nairne, 1991).
The final model that I discuss is my own feature model (Nairne, 1988, 1990, 2001; Neath & Nairne, 1995). All forgetting in the feature model is attributed to interference, either from feature overwriting or from incorrect interpretation of the primary memory trace. Rehearsal can play a role in the model, as a mechanism for effectively re-presenting list items, but rehearsal plays no real role in producing standard immediate memory phenomena such as the word length effect or even the effects of articulatory suppression on performance (see also Neath, 2000). The model assumes that residual remnants of perceptual processing remain in primary memory after list presentation. These primary memory traces are represented as vectors of features and can be overwritten, based on similarity, by subsequent list items. At the point of recall, surviving traces exist in a degraded or blurry form and must be interpreted prior to recall. Most of the interesting effects of immediate retention arise out of the interpretation process.
The feature model has been applied successfully to most of the standard phenomena of immediate memory, including the modality and suffix effects. One of its most important assumptions is the idea that the trace interpretation process is guided by the presence or absence of distinctive features. Correct performance hinges on the presence of features in the degraded trace that uniquely specify one of the possible recall candidates. To the extent that primary memory traces contain features that are matched in all of the presented items, such as a common sound or phoneme, performance suffers. It is this characteristic of the model that explains the phonological similarity effect, as well as long-standing phenomena such as the von Restorff effect (see Kelley & Nairne, 2001). More importantly, the trace interpretation process is assumed to resemble the kinds of cue-driven retrieval processes that guide all forms of remembering, regardless of the time scales involved (see Nairne, 2002). The feature model, like the other unitary models discussed, assigns no special mnemonic laws or properties (such as decay) to remembering over the short term.
Conclusions
Transient memories, discussed here in the form of sensory and short-term memory, clearly serve highly adaptive functions in human cognitive processing. Sensory memories enable us to prolong the present, for the briefest of intervals; short-term memories comprise the ingredients of conscious awareness and play a vital role, among other things, in the comprehension and production of spoken language. Compared to the study of long-term retention, studying transient memories is a relatively recent enterprise, commencing with full vigor only in the second half of the twentieth century. As we have seen, many issues remain unresolved, and fundamental controversies continue. Does short-term retention follow its own unique operating laws? Is it necessary to propose processes, such as decay, that apply uniquely to remembering over the short term? Is sensory persistence truly an evolved form of remembering, serving its own special function, or is it simply an artifact of the properties of neural networks?
Despite these controversies, few questions remain about the data to be explained. In the presence of distractor activity, we can still remember only a handful of unrelated items for more than 10 or 20 s; when an array of unrelated letters is briefly flashed, a partial reporting of the array is still dramatically better than reporting of the whole. Whatever form the final accounting of these phenomena takes, it will undoubtedly provide insight into far more than simply remembering over the short term. The study of transient memories is likely to provide a clear window for understanding all forms of memory, both short and long.
Bibliography:
- Anderson, J. R., & Matessa, M. (1997). A production system theory of serial memory. Psychological Review, 104, 728–774.
- Atkinson, R. C., & Shiffrin, R. M. (1968). Human memory: A proposed system and its control processes. In K. W. Spence & J. T. Spence (Eds.), The psychology of learning and motivation (Vol. 2, pp. 89–105). New York: Academic Press.
- Averbach, E., & Coriell, H. S. (1961). Short-term memory in vision. Bell Systems Technical Journal, 40, 309–328.
- Ayres, T. J., Jonides, J., Reitman, J. S., Egan, J. C., & Howard, D. A. (1979). Differing suffix effects for the same physical suffix. Journal of Experimental Psychology: Human Learning and Memory, 5, 315–321.
- Baddeley, A. D. (1986). Working memory. Oxford, UK: Oxford University Press.
- Baddeley, A. D. (1996). Exploring the central executive. Quarterly Journal of Experimental Psychology, 49A, 5–28.
- Baddeley, A. D. (2000). The episodic buffer: A new component of working memory? Trends in Cognitive Sciences, 4, 417–423.
- Baddeley, A. D., Gathercole, S. E., & Papagno, C. (1998). The phonological loop as a language learning device. Psychological Review, 105, 158–173.
- Baddeley, A. D., & Hitch. G. (1974). Working memory. In G. H. Bower (Ed.), The psychology of learning and motivation (Vol. 8, pp. 47–89). New York: Academic Press.
- Baddeley, A. D., Lewis, V. J., & Vallar, G. (1984). Exploring the articulatory loop. Quarterly Journal of Experimental Psychology, 36, 233–252.
- Baddeley, A. D., & Lieberman, K. (1980). Spatial working memory. In R. Nickerson (Ed.), Attention and performance VIII (pp. 521–539). Hillsdale, NJ: Erlbaum.
- Baddeley, A. D., & Logie, R. H. (1999). Working memory: The multicomponent model. In A. Miyake & P. Shah (Eds.), Models of working memory (pp. 28–61). Cambridge, UK: Cambridge University Press.
- Baddeley, A. D., & Scott, D. (1971). Short term forgetting in the absence of proactive interference. Quarterly Journal of Experimental Psychology, 23, 275–283.
- Baddeley, A. D., Thomson, N., & Buchanan, M. (1975). Word length and the structure of short-term memory. Journal of Verbal Learning and Verbal Behavior, 14, 575–589.
- Bjork, R. A. (2001). Recency and recovery in human memory. In L. Roediger, J. S. Nairne, I. Neath, & A. Surprenant (Eds.), The nature of remembering: Essays in honor of Robert G. Crowder (pp. 211–232). Washington, DC: American Psychological Association.
- Bowen, R. W., Pola, J., & Matin, L. (1974). Visual persistence: Effects of flash luminance, duration and energy. Vision Research, 14, 295–303.
- Broadbent, D. E. (1958). Perception and communication. London: Pergamon Press.
- Brown, G. D. A., McCormack, T., & Chater, N. (2001). The chronological organisation of memory: Common psychological foundations for remembering and time. In C. Hoerl & T. McCormack (Eds.), Time and memory: Issues in philosophy and psychology. Oxford, UK: Oxford University Press.
- Brown, G. D. A., Preece, T., & Hulme, C. (2000). Oscillator-based memory for serial order. Psychological Review, 107, 127–181.
- Brown, J. (1958). Some tests of the decay theory of immediate memory. Quarterly Journal of Experimental Psychology, 10, 12–21.
- Burgess, N., & Hitch, G. J. (1999). Memory for serial order: A network model of the phonological loop and its timing. Psychological Review, 106, 551–581.
- Campbell, R., & Dodd, B. (1980). Hearing by eye. Quarterly Journal of Experimental Psychology, 32, 85–99.
- Caplan, D., Rochon, E., & Waters, G. S. (1992). Articulatory and phonological determinants of word-length effects in span tasks. Quarterly Journal of Experimental Psychology, 45A, 177–192.
- Cofer, C. N., & Davidson, E. H. (1968). Proactive interference in STM for consonant units of two sizes. Journal of Verbal Learning and Verbal Behavior, 7, 268–270.
- Coltheart, M. (1980). Iconic memory and visual persistence. Perception & Psychophysics, 27, 183–228.
- Conrad, R. (1964). Acoustic confusion in immediate memory. British Journal of Psychology, 55, 75–84.
- Corballis, M. C. (1966). Rehearsal and decay in immediate recall of visually and aurally presented items. Canadian Journal of Psychology, 20, 43–51.
- Cowan, N. (1984). On short and long auditory stores. Psychological Bulletin, 96, 341–370.
- Cowan, N. (1988). Evolving conceptions of memory storage, selective attention, and their mutual constraints within the human informationprocessingsystem.PsychologicalBulletin,104,163– 191.
- Cowan, N. (1995). Attention and memory: An integrated framework. New York: Oxford University Press.
- Cowan, N. (2001). The magical number 4 in short-term memory: A reconsideration of mental storage capacity. Behavioral and Brain Sciences, 24, 87–185.
- Crowder, R. G. (1976). Principles of learning and memory. Hillsdale, NJ: Erlbaum.
- Crowder, R. G. (1978). Memory for phonologically uniform lists. Journal of Verbal Learning and Verbal Behavior, 17, 73–89.
- Crowder, R. G., & Morton, J. (1969). Precategorical acoustic storage (PAS). Perception & Psychophysics, 5, 365–373.
- Crowder, R. G., & Surprenant, A. M. (2000). Sensory memory. In A. E. Kazdin (Ed.), Encyclopedia of psychology (Vol. 7, pp. 227–229). New York: Oxford University Press and American Psychological Association.
- Daneman, M., & Carpenter, P. A. (1980). Individual differences in working memory and reading. Journal of Verbal Learning and Verbal Behavior, 19, 450–466.
- Darwin, C. J. (1976). The perception of speech. In E. C. Carterette & M. P. Friedman (Eds.), Handbook of perception (Vol. 7, pp. 175–218). New York:Academic Press.
- Darwin, C. J., Turvey, M. T., & Crowder, R. G. (1973). An auditory analogue of the Sperling partial-report procedure. Cognitive Psychology, 3, 255–267.
- Dempster, F. N. (1992). The rise and fall of the inhibitory mechanism: Toward a unified theory of cognitive development and aging. Developmental Review, 12, 45–75.
- Dillon, R. F., & Bittner, L. A. (1975). Analysis of retrieval cues and release from proactive inhibition. Journal of Verbal Learning and Verbal Behavior, 14, 616–622.
- Di Lollo, V., Hogben, J. H., & Dixon, P. (1994). Temporal integration and segregation of brief visual stimuli: Patterns of correlation in time. Perception & Psychophysics, 55, 373–386.
- Dixon, , Gordon, R. D., Leung, A., & DiLollo,V.(1997). Attentional components of partial report. Journal of Experimental Psychology: Human Perception and Performance, 23, 1253–1271.
- Dosher, A., & Ma, J.-J. (1998).Output lossor rehearsalloop?Output-time versus pronunciation-time limits in immediate recall for forgetting-matched materials. Journal of Experimental Psychology: Learning, Memory, and Cognition, 24, 316–335.
- Dosher, B. A., & McElree, B. (1992). Memory search: Retrieval processes in short-term and long-term retention. In L. R. Squire (Ed.), Encyclopedia of learning and memory (pp. 398–405). New York: Macmillan.
- Ebbinghaus, H. (1964). Memory: A contribution to experimental psychology. New York: Dover. (Original work published 1885)
- Efron, R. (1970). Effect of stimulus duration on perceptual onset and offset latencies. Perception & Psychophysics, 8, 231–234.
- Ellis, N. C., & Hennelley, R. A. (1980). A bilingual word-length effect: Implications for intelligence testing and the relative ease of mental calculation in Welsh and English. British Journal of Psychology, 71, 43–52.
- Engle, R. E., Kane, M. J., & Tuholski, S. W. (1999). Individual differences in working memory capacity and what they tell us about controlled attention, general fluid intelligence, and functions of the prefrontal cortex. In A. Miyake & P. Shah (Eds.), Models of working memory (pp. 102–134). Cambridge, UK: Cambridge University Press.
- Eriksen, C. W., & Collins, J. F. (1967). Some temporal characteristics of visual pattern perception. Journal of Experimental Psychology, 74, 476–484.
- Estes, W. K. (1972). An associative basis for coding and organization in memory. In A. W. Martin & E. Martin (Eds.), Coding processes in human memory (pp. 161–190). Washington, DC: Winston.
- Estes, W. K. (1991). On types of item coding and source of recall in short-term memory. In W. E. Hockley & S. Lewandowsky (Eds.), Relating theory and data: Essays on human memory in honor of Bennet B. Murdock (pp. 155–174). Hillsdale, NJ: Erlbaum.
- Estes, W. K. (1997). Processes of memory loss, recovery, and distortion. Psychological Review, 104, 148–169.
- Francis, G. (1999). Spatial frequency and visual persistence: Cortical reset. Spatial Vision, 12, 31–50.
- Gardiner, J. M., Craik, F. I. M., & Birtwistle, J. (1972). Retrieval cues and release from proactive inhibition. Journal of Verbal Learning and Verbal Behavior, 11, 778–783.
- Greene, R. L. (1992). Human memory: Paradigms and paradoxes. Hillsdale, NJ: Erlbaum.
- Greene, R. L. (1996). The influence of experimental design: The example of the Brown-Peterson paradigm. Canadian Journal of Experimental Psychology, 50, 240–242.
- Greene, R. L. & Crowder, R. G. (1984). Modality and suffix effects in the absence of auditory stimulation. Journal of Verbal Learning and Verbal Behavior, 23, 371–382.
- Grossberg, S. (1994). 3-D vision and figure-ground separation by the visual cortex. Perception & Psychophysics, 55, 48–120.
- Haber, R. N. (1983). The impending demise of the icon: A critique of the concept of iconic storage in visual information processing. Behavioral and Brain Sciences, 6, 1–10.
- Halford, G. S., Mayberry, M. T., & Bain, J. D. (1988). Set-size effects in primary memory: An age-related capacity limitation? Memory & Cognition, 16, 480–487.
- Healy, A. F. (1974). Separating item from order information in short-term memory. Journal of Verbal Learning and Verbal Behavior, 13, 644–655.
- Healy, A. F., & Cunningham, T. F. (1995). Very rapid forgetting: A reply to Muter. Memory & Cognition, 23, 387–392.
- Healy, A. F., & McNamara, D. S. (1996). Verbal learning and memory: Does the modal model still work? Annual Review of Psychology, 47, 143–172.
- Henson, R. N. A. (1996). Short-term memory for serial order. Unpublished doctoral dissertation, University of Cambridge, Cambridge, UK.
- Henson, R. N. A. (1998). Short-term memory for serial order: The start-end model. Cognitive Psychology, 36, 73–137.
- Henson, R. N. A. (1999). Positional information in short-term memory: Relative or absolute? Memory & Cognition, 27, 915–927.
- Hitch, G. J., Burgess, N., Towse, J. N., & Culpin, V. (1996). Temporal grouping effects in immediate recall: A working memory analysis. Quarterly Journal of Experimental Psychology, 49A, 116–139.
- Hulme, C., Maughan, S., & Brown, G. D. A. (1991). Memory for familiar and unfamiliar words: Evidence for a long-term memory contribution to short-term memory span. Journal of Memory and Language, 30, 685–701.
- Hulme, C., Roodenrys, S., Schweickert, R., Brown, G. D. A., Martin, S., & Stuart, G. (1997). Word frequency effects on shortterm memory tasks: Evidence for a multinomial processing tree model of immediate serial recall. Journal of Experimental Psychology: Learning, Memory, and Cognition, 23, 1217–1232.
- Hulme, C., Thomson, N., Muir, C., & Lawrence, A. (1984). Speech rate and the development of short-term memory span. Journal of Experimental Child Psychology, 38, 241–253.
- Hulme, C., & Tordoff, V. (1989). Working memory development: The effects of speech rate, word length, and acoustic similarity on serial recall. Journal of Experimental Child Psychology, 47, 72–87.
- Kane, M. J., & Engle, R. W. (2000). Working-memory capacity, proactive interference, and divided attention: Limits on longterm memory retrieval. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 336–358.
- Kelley, M. R., & Nairne, J. S. (2001). von Restorff revisited: Isolation, generation, and memory for order. Journal of Experimental Psychology: Learning, Memory, and Cognition, 27, 54–66.
- Keppel, G., & Underwood, B. J. (1962). Proactive inhibition in short-term retention of single items. Journal of Verbal Learning and Verbal Behavior, 1, 153–161.
- Lee, C. L., & Estes, W. K. (1981). Item and order information in short-term memory: Evidence for multilevel perturbation processes. Journal of Experimental Psychology: Human Learning and Memory, 7, 149–169.
- Loftus, G. R., & Irwin, D. E. (1998). On the relations among different measures of visible and informational persistence. Cognitive Psychology, 35, 135–199.
- Logie, R. H. (1995). Visuo-spatial working memory. Hove, UK: Erlbaum.
- Lovatt, P., Avons, S. E., & Masterson, J. (2000). The word-length effect and disyllabic words. Quarterly Journal of Experimental Psychology, 53A, 1–22.
- Massaro, D. W. (1970). Perceptual processes and forgetting in memory tasks. Psychological Review, 77, 557–567.
- Massaro, D. W. (1975). Experimental psychology and information processing. Chicago: Rand-McNally.
- Massaro, D. W., & Loftus, G. R. (1996) Sensory and perceptual storage. In E. L. Bjork & R. A. Bjork (Eds.), Handbook of perception and cognition (Vol. 10, pp. 67–99). New York: Academic Press.
- McElree, B. (1998). Attended and non-attended states in working memory: Accessing categorized structures. Journal of Memory and Language, 38, 225–252.
- McElree, B., & Dosher, B. A. (1989). Serial position and set size in short-term memory: Time course of recognition. Journal of Experimental Psychology: General, 118, 346–373.
- McElree, B., & Dosher, B. A. (1993). Serial retrieval processes in the recovery of order information. Journal of Experimental Psychology: General, 122, 291–315.
- McGeoch, J. A. (1932). Forgetting and the law of disuse. Psychological Review, 39, 352–370.
- Melton, A. W. (1963). Implications of short-term memory for a general theory of memory. Journal of Verbal Learning and Verbal Behavior, 2, 1–21.
- Mewhort, D. J. K., Campbell,A. J., Marchetti, F. M., & Campbell, J. I.D.(1981).Identification,localization,and“iconicmemory:”An evaluation of the bar-probe task. Memory & Cognition, 9, 50–67.
- Miller, G. A. (1956). The magical number seven plus or minus two: Some limits on our capacity for processing information. Psychological Review, 63, 81–97.
- Miyake, A., & Shah, P. (1999). Models of working memory. Cambridge, UK: Cambridge University Press.
- Morton, J., Crowder, R. G., & Prussin, H. A. (1971). Experiments with the stimulus suffix effect. Journal of Experimental Psychology, 91, 169–190.
- Morton, N., & Morris, R. G. (1995). Image transformation dissociated from visuospatial working memory. Cognitive Neuropsychology, 12, 767–791.
- Murdock, B. B., Jr. (1961). The retention of individual items. Journal of Experimental Psychology, 62, 618–625.
- Murray, D. J. (1968) Articulation and acoustic confusability in short-term memory. Journal of Experimental Psychology, 78, 679–684.
- Muter, P. (1980). Very rapid forgetting. Memory & Cognition, 8, 174–179.
- Näätänen, R., & Winkler, I. (1999). The concept of auditory stimulus representation in cognitive neuroscience. Psychological Bulletin, 125, 826–859.
- Nairne, J. S. (1988). A framework for interpreting recency effects in immediate serial recall. Memory & Cognition, 16, 343–352.
- Nairne, J. S. (1990). A feature model of immediate memory. Memory & Cognition, 18, 251–269.
- Nairne, J. S. (1991). Positional uncertainty in long-term memory. Memory & Cognition, 19, 332–340.
- Nairne, J. S. (1992). The loss of positional certainty in long-term memory. Psychological Science, 3, 199–202.
- Nairne, J. S. (1996). Short-term/working memory. In E. L. Bjork & R. A. Bjork (Eds.), Memory (pp. 101–126). New York: Academic Press.
- Nairne, J. S. (2001). A functional analysis of primary memory. In H. L. Roediger, J. S. Nairne, I. Neath, & A. M. Surprenant (Eds.), The nature of remembering: Essays in honor of Robert G. Crowder (pp. 282–296). Washington, DC: American Psychological Association.
- Nairne, J. S. (2002). Remembering over the short-term: The case against the standard model. Annual Review of Psychology, 53, 53–81.
- Nairne, J. S., & Walters, V. L. (1983). Silent mouthing produces modality- and suffix-like effects. Journal of Verbal Learning and Verbal Behavior, 22, 475–483.
- Naveh-Benjamin, M., & Ayres, T. J. (1986). Digit span, reading rate, and linguistic relativity. Quarterly Journal of Experimental Psychology, 38A, 739–751.
- Neath, I. (1998). Human memory: An introduction to research, data, and theory. Pacific Grove, CA: Brooks/Cole.
- Neath, I. (2000). Modelling the effects of irrelevant speech on memory. Psychonomic Bulletin & Review, 7, 403–423.
- Neath, I., & Crowder, R. G. (1990). Schedules of presentation and temporal distinctiveness in human memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16, 316– 327.
- Neath, I., Kelley, M. R., & Surprenant, A. M. (2001). Fill-in and infill errors in immediate serial recall. Manuscript submitted for publication.
- Neath, I., & Knoedler, A. J. (1994). Distinctiveness and serial position effects in recognition and sentence processing. Journal of Memory and Language, 33, 776–795.
- Neath, I., & Nairne, J. S. (1995). Word-length effects in immediate memory: Overwriting trace-decay theory. Psychonomic Bulletin & Review, 2, 429–441.
- Neath, I., Surprenant, A. M., & Crowder, R. G. (1993). The context dependent stimulus suffix effect. Journal of Experimental Psychology: Learning, Memory, and Cognition, 19, 698–703.
- Osgood, C. E. (1953). Method and theory in experimental psychology. New York: Oxford University Press.
- Page, M. P. A., & Norris, D. (1998). The primacy model: A new model of immediate serial recall. Psychological Review, 105, 761–781.
- Papagno, C., Valentine, T., & Baddeley, A. D. (1991). Phonological short-term memory and foreign-language vocabulary learning. Journal of Memory and Language, 30, 331–347.
- Peterson, L. R., & Peterson, M. J. (1959). Short-term retention of individual verbal items. Journal of Experimental Psychology, 58, 193–198.
- Proctor, R. W., & Fagnani, C. A. (1978). Effects of distractorstimulus modality in the Brown-Peterson distractor task. Journal of Experimental Psychology: Human Learning and Memory, 4, 676–684.
- Quinn, J. G., & McConnell, J. (1996). Irrelevant pictures in visual working memory. Quarterly Journal of Experimental Psychology, 49A, 200–215.
- Ratcliff, R. (1978). A theory of memory retrieval. Psychological Review, 85, 59–108.
- Reed, A. V. (1976). The time course of recognition in human memory. Memory & Cognition, 4, 16–30.
- Roodenrys, S., Hulme, C., Alban, J., & Ellis, A. W. (1994). Effects of word frequency and age of acquisition on short-term memory span. Memory & Cognition, 22, 695–701.
- Rosen, V. M., & Engle, R. W. (1998). Working memory capacity and suppression. Journal of Memory and Language, 39, 418–436.
- Ryan, J. (1969). Grouping and short-term memory: Different means and patterns of groups. Quarterly Journal of Experimental Psychology, 21, 137–147.
- Schweickert, R. (1993). A multinomial processing tree model for degradation and redintegration in immediate recall. Memory & Cognition, 21, 167–175.
- Schweickert, R., & Boruff, B. (1986). Short-term memory capacity: Magic number or magic spell? Journal of Experimental Psychology: Learning, Memory, and Cognition, 12, 419–425.
- Schweickert, R., Guentert, L., & Hersberger, L. (1990). Phonological similarity, pronunciation rate, and memory span. Psychological Science, 1, 74–77.
- Sebrechts, M. M., Marsh, R. L., & Seamon, J. G. (1989). Secondary memoryandveryrapidforgetting.Memory&Cognition,17,693– 700.
- Service,E.(1998).Theeffectofwordlengthonimmediateserialrecall depends on phonological complexity, not articulatory duration. Quarterly Journal of Experimental Psychology, 51A, 283–304.
- Shallice, T., & Warrington, E. K. (1970). Independent functioning of verbal memory stores: A neuropsychological study. Quarterly Journal of Experimental Psychology, 22, 261–273.
- Shiffrin, R. M. (1999). 30 years of memory. In C. Izawa (Ed.), On human memory: Evolution, progress, and reflections on the 30th anniversaryoftheAtkinson-Shiffrinmodel(pp.17–33).Mahwah, NJ:Erlbaum.
- Slamecka, N. J. (1967). Human memory and learning: Selected readings. New York: Oxford University Press.
- Sperling, G. (1960). The information available in brief visual presentation. Psychological Monographs, 74, 1–29.
- Sternberg, S. (1966). High-speed scanning in human memory. Science, 153, 652–654.
- Tan, L., & Ward, G. (2000). Arecency-based account of the primacy effect in free recall. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 1589–1625.
- Tehan, G., & Humphreys, M. S. (1995). Transient phonemic codes and immunity to proactive interference. Memory & Cognition, 23, 181–191.
- Tehan, G., & Humphreys, M. S. (1996). Cuing effects in short-term recall. Memory & Cognition, 24, 719–732.
- Tehan, G., & Humphreys, M. S. (1998). Creating proactive interference in immediate recall: Building a dog from a dart, a mop and a fig. Memory & Cognition, 26, 477–489.
- Tehan, G., & Lalor D. M. (2000). Individual differences in memory span: The contribution of rehearsal, access to lexical memory and output speed. Quarterly Journal of Experimental Psychology, 53A, 1012–1038.
- Townsend, T., & Ashby, F. G. (1983). The stochastic modeling of elementary psychological processes. Cambridge, UK: Cambridge University Press.
- Tulving, E. (1983). Elements of episodic memory. New York: Oxford University Press.
- Turner, L., & Engle, R. W. (1989). Is working memory capacity task dependent? Journal of Memory and Language, 28, 127–154.
- Turvey, M. T. (1973). On peripheral and central processes in vision: Inferences from an information processing analysis of masking with patterned stimuli. Psychological Review, 80, 1–52.
- Turvey, M. T., Brick, P., & Osborn, J. (1970). Proactive interference in short-term memory as a function of prior-item retention interval. Quarterly Journal of Experimental Psychology, 22, 142–147.
- Walker, I., & Hulme, C. (1999). Concrete words are easier to recall than abstract: Evidence for a semantic contribution to short-term serial recall. Journal of Experimental Psychology: Learning, Memory, & Cognition, 25, 1256–1271.
- Watkins, O. C., & Watkins, M. J. (1975). Buildup of proactive inhibition as a cue-overload effect. Journal of Experimental Psychology: Human Learning and Memory, 104, 442–452.
- Watkins, O. C., & Watkins, M. J. (1980). The modality effect and echoic persistence. Journal of Experimental Psychology: General, 109, 251–278.
- Wickelgren, W. A. (1967). Rehearsal grouping and hierarchical organization of serial position cues in short-term memory. Quarterly Journal of Experimental Psychology, 19, 97–102.
- Wickelgren, W. A., Corbett, A. T., & Dosher, B. A. (1980). Priming and retrieval from short-term memory: A speed-accuracy tradeoff analysis. Journal of Verbal Learning and Verbal Behavior, 19, 387–404.
- Wickens, D. D. (1970). Encoding categories of words: An empirical approach to meaning. Psychological Review, 77, 1–15.
- Wickens, D. D., Moody, M. J., & Dow, R. (1981). The nature and timing of the retrieval process and of interference effects. Journal of Experimental Psychology: General, 110, 1–20.
- Wright, A. A., Santiago, H. C., Sands, S. F., Kendrick, D. F., & Cook, R. G. (1985). Memory processing of serial lists by pigeons, monkeys, and people. Science, 229, 287–28
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality