
Sample Action Selection Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. iResearchNet offers academic assignment help for students all over the world: writing from scratch, editing, proofreading, problem solving, from essays to dissertations, from humanities to STEM. We offer full confidentiality, safe payment, originality, and money-back guarantee. Secure your academic success with our risk-free services.
Action selection refers to how a decision is made, typically under speeded response conditions, regarding which of two or more actions to take in response to perceptual events. It is usually studied using choice-reaction tasks in which subjects make assigned responses to stimuli as quickly and accurately as possible, and reaction time (RT) and response accuracy are measured. Action selection is often called response selection, but the term action selection has come to be used more frequently in recent years to emphasize that responses in choicereaction tasks are goal-directed actions (Prinz, 1997).
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
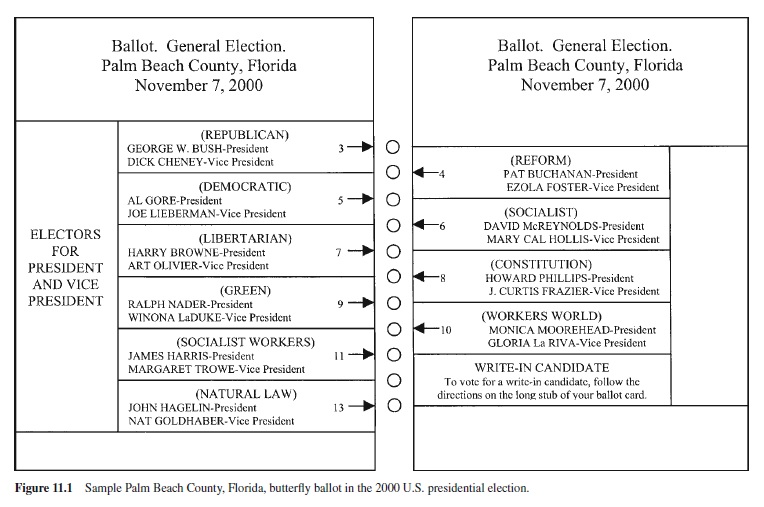
A recent example of the importance of action selection concerns the notorious butterfly ballot used in Palm Beach County, Florida, for the 2000 U.S. presidential election. The ballot, shown in Figure 11.1, listed the names of candidates in two columns, with the appropriate response being to insert a stylus into a punch hole assigned to the candidate of choice among a centered column of holes. Although there was no fixed time limit for responding, the voters’ task was speeded in the sense that a limited number of voting booths were available, with many voters needing to use them. With this ballot, some voters apparently selected the second punch hole on the list, voting for Pat Buchanan, rather than the third punch hole, which was assigned to Al Gore, for whom they intended to vote. This selection error occurred because Gore was listed in the second position of the left-hand column, immediately below the major opposing candidate, George W. Bush. Punch ballots most often list all candidates on the lefthand side, and their corresponding punch holes in the same order on the right. Because the relative location of Gore’s position in the left-hand candidate list was second, previous experience would lead voters to expect that the second hole should be punched to vote for him. Moreover, this expectancy is consistent with the general principle that people tend to make the response whose relative location corresponds to that of the stimulus. Consequently, it is not surprising that some voters would incorrectly punch the second hole instead of the third one, even though arrows were used to mark the designated punch holes for the candidates. The poor design of the ballot caused a sufficient number of unintended votes for Buchanan, as well as discarded ballots for which the second and third holes were punched, costing Gore the election.
As this example illustrates, the topic of action selection is undoubtedly important. However, action selection tends to be viewed as peripheral to mainstream cognitive psychology in the United States, as reflected in the fact that the topic is rarely mentioned in undergraduate cognitive psychology texts. The view of many cognitive psychologists seems to be that input and central processes can be investigated without one’s having to be concerned with the translation of the outcome of these processes into output. This view is ironic, given that a major impetus to the rise of contemporary cognitive psychology was research on human performance conducted by Paul Fitts (see Fitts & Posner, 1967), Donald Broadbent (1958), and others in the 1950s. Outside of the United States, more recognition has been given to the importance of selection and execution of action in human information processing. Action selection is seen as fundamental because it involves the interface between perception and action. It is the theme of this research paper that action selection is of vital importance to many of the phenomena studied in contemporary cognitive psychology.
Fundamental Issues, Models, and Theories
Historical Background
Astronomers in the first half of the nineteenth century made the initial contribution to the measurement of RT by estimating the time it took a star to reach the midline of a grid of vertical lines relative to when it first entered the grid (see Woodworth, 1938). Although this was a clever method of measuring RT, individual differences in the judgment of when the star entered and reached the midline resulted in unreliable readings from one astronomer to another. In an attempt to compensate for individual differences, a personal equation was developed in which a constant correction was made in order to equate the readings of astronomers. However, later investigations showed that the difference between two individuals was not constant after all.
The study of action selection was of central concern in the last half of the nineteenth century. Interest arose out of issues concerning the speed of nerve transmission. Most physiologists thought that nerve transmission occurred too rapidly to be measured. However, Helmholtz (1850) conducted an experiment in which he stimulated motor nerves of frogs and measured the time between the presentation of the stimulus and muscular contraction. He estimated the rate of nerve transmission to be 26 m/s. One important contribution of this work was to demonstrate that the durations of nervous systems’ processes are measurable. Helmholtz was also the first to measure RT in a procedure intended to calculate the speed of nerve transmission in humans. This procedure involved measuring RT as a function of the distance away from the brain by applying a shock to the skin. However, Helmholtz concluded that this procedure does not yield an accurate measure of nerve conduction because the measurements “suffer from the unfortunate fact that a part of the measured time depends on mental processes” (Helmholtz, 1867, p. 228).
The research of Helmholtz and others using RT to estimate the speed of nerve conduction stimulated Donders and his students to pursue the use of RT as a means for measuring mental processes. De Jagger’s dissertation (1865/1970) provided the first account of the experiments conducted in Donders’s lab. The first part of De Jagger’s study continued Helmholtz’s notion of measuring the speed of nerve conduction, but the second part focused on measuring the time required to identify a stimulus and select a motor response. In one set of experiments, subjects were required to respond to a red light with the right hand and a white light with the left hand. The mean RT was 356 ms, which was 172 ms longer than a simple reaction (executing a single response when a stimulus is presented) to the same stimuli. De Jagger interpreted this time as the duration of the central processes involving stimulus discrimination and response initiation.
Donders (1868/1969) formalized the subtractive method used by De Jagger, emphasizing specifically that the time for a particular process could be estimated by adding that process to a task and taking the difference in RT between the two tasks. He distinguished three types of reactions: type a (simple reaction), type b (choice reaction), and type c (go or no-go reaction; responding to one stimulus but not another). These types of reactions allowed separate measures of the stimulus identification and decision processes that were assessed together by De Jagger. The difference between the type-c and type-a reactions was presumed to reflect the time for stimulus identification, and the difference between the type-b and type-c reactions the time for “expression of the will” (p. 424).
Reaction time research in general, and the study of action selection in particular, continued to flourish throughout the remainder of the nineteenth century (see Jastrow, 1890). Wundt (1883) criticized Donders for using the type-c reaction as a measure of stimulus identification, reasoning that subjects must distinguish whether to respond, and suggested using the type-d reaction instead as a pure measure. The typed reaction is measured by presenting subjects with the same stimuli and having them make the same response every time, as in the type-a reaction, with the difference being that they are instructed not to respond until they have identified the stimulus. However, Wundt’s type-d reaction quickly fell out of favor because it is subjective and highly variable, and after practice, the type-d reaction time does not differ from the type-a reaction time. Criticisms of the subtractive method in general led to its demise in the early twentieth century.
Methodological and Modeling Issues
With the advent of the information processing approach in the 1950s and 1960s, the subtractive method was resurrected. This method, and the stage analysis of RT data on which it is based, came to be seen as sufficiently important to establish Donders as a major figure in the history of human performance. One influential use of the subtractive method was to estimate the rate of mental rotation by varying the amount that one stimulus was rotated relative to another to which it was to be compared, and measuring the slope of the RT function (Cooper & Shepard, 1973). Mean RT increased by approximately 240 ms for each 20° increase in angle of rotation, suggesting a continuous transformation in which each degree of rotation took about 12 ms.
A major advance in stage analysis of RT data was the development of the additive factors method by Sternberg (1969). Like the subtractive method, the additive factors method assumes discrete serial processing stages. However, whereas the subtractive method provides duration estimates for assumed stages, the additive factors method provides a way to discover the stages themselves. Sternberg showed that if two or more factors each influence the durations of distinct stages, then the effect of one of the factors on total duration will be invariant across the levels of the other factors: That is, the effects of the variables on RT will be additive. If two factors have interactive effects on RT, then they must influence at least one common stage. Thus, Sternberg advocated the use of multifactor experiments in which the presence or absence of interactions among variables is used to determine the processing stages involved in task performance.
Numerous limitations of the additive factors method have been enunciated, including problems of accepting the null hypothesis for additivity, assuming serial processing stages with no feedback loops, and assuming constant output from each stage (see Pachella, 1974). Despite these limitations, the method has proven to be a useful tool for analyzing the structure of information processing in a variety of tasks (see Sternberg, 1998) because, as Sanders (1998) states, “the method appears to provide a successful summary of a large amount of experimental data” (p. 65). One criterion for evaluating the additive factors method is stage robustness: The relations between two factors should not change as a function of levels of other factors. Although there are exceptions, stage robustness has generally been found to hold (Sanders, 1998).
Discrete and Continuous Models of Information Processing
Sternberg’s (1969) additive factors method is based on a view of human information processing that assumes that the processing sequence between stimulus and response consists of a series of discrete stages, with each stage completing its processing before the next stage begins (see Figure 11.2, left side). Other models allow for parallel or overlapping operation of the different processing stages. McClelland (1979) proposed the cascade model of information processing in which partial information at one subprocess, or stage, is transferred to the next (see Figure 11.2, right side). The model assumes that each stage is continuously active and its output is a continuous value that is always available to the next stage. As in the discrete stage model, it is also assumed that each stage operates only on the output from the preceding stage. The output of the final stage indicates which of the alternative responses to execute.

In the cascade model, an experimental manipulation may affect a stage by altering the rate of activation or the asymptotic level of activation. The asymptotic level is equivalent to the stage output in the discrete stage model, which is assumed to be constant, and the activation rate determines the speed at which the final output is attained. Although the assumptions of the cascade model are different from those of the discrete stage model from which the additive factors method was derived, the patterns of interactivity and additivity can be interpreted similarly. For the cascade model, if two variables affect the rate parameter of the same stage, their effects on RT will be interactive; if each variable affects the rate parameter of a different stage, their effects on RT will be additive. In sum, as long as it is assumed that the final output of a stage does not vary as a function of the manipulations, then use of the additive factors logic to interpret the RT patterns does not require an assumption of discrete stages.
Miller (1988) argued that the discrete versus continuous categorization should not be viewed as dichotomous but as extremes on a quantitative dimension called grain size. In his words, “a variable is more continuous to the extent that it has a small grain size and more discrete to the extent that it has a large one” (p. 195). Miller suggested that there are three different senses in which models of human information processing can be characterized as discrete or continuous: representation, transformation, and transmission.
Representation refers to the discrete/continuous nature of the input and output codes for the processing stage. For example, if the locations of stimuli and responses in two-choice spatial reaction tasks are coded as left or right in terms of relative position, as is often assumed, the spatial codes are discrete. However, if the locations are represented in terms of absolute positions in physical space, then the representations are continuous. Transformation refers to the nature of the operation that the processing stage performs. The transformation of the stage that performs mental rotation is typically characterized as continuous, in the sense that the mentally rotated object passes through a continuum of intermediate states from its initial orientation to the final orientation (Cooper & Shepard, 1973).Acompletely discrete transformation would be to generate the final orientation in a single step. For transmission, a model is discrete if the processing of successive stages cannot have temporal overlap; that is, the next stage in the sequence must wait until processing of the immediately preceding stage is completed. The discrete stage model underlying Sternberg’s (1969) additive factorsmethodpostulates discrete representation and transmission. McClelland’s (1979) cascade model, on the other hand, postulates continuous representation and transmission, as well as transformation.
A variety of models exist that are intermediate to these two extremes. One such model is Miller’s (1982, 1988) asynchronous discrete coding model. This model assumes that most stimuli are composed of features, and these features are identified separately. The processing is discrete in that each feature must be identified before output about it can be passed to the response-selection stage. However, the identity of one feature may be passed to response selection while stimulus identification processes are still operating on other features.
Speed-Accuracy Trade-Off
The subtractive and additive factors methods are usually based solely on RT data. However, RT in any specific task situation is related to the number of errors that one is willing to make. Aperson can respond rapidly and make many errors or slowly and make few errors. This relation is called the speedaccuracy trade-off, and the function plotting speed versus accuracy is known as the speed-accuracy operating characteristic. For RT research, two aspects are crucial. First, if slower RT is accompanied by lower error rate, then the RT difference cannot be attributed unambiguously to differences in processing efficiency. Second, under conditions in which accuracy is relatively high, as in most choice-reaction studies, a small difference in error rate can translate into a large difference in RT.
Because of this close relation between speed and accuracy, some researchers have advocated conducting experiments in which the speed-accuracy criterion is varied between blocks of trials (Dosher, 1979; Pachella, 1974). There are numerous ways to vary the speed-accuracy criterion: payoffs, instructions, deadlines, time bands (responding within a certain time interval), and response signals (responding when the response signal is presented; see Wickelgren, 1977, for details). When a speed-accuracy function is obtained, information is provided about the intercept (time at which accuracy exceeds chance), asymptote (the maximal accuracy), and rate of ascension from the intercept to the asymptote, each of which may reflect different processes. Thus, a speedaccuracy study has the potential to be more informative than one based solely on RT. However, speed-accuracy studies require 5–10 times more data than RT studies and, in many circumstances, do not provide better insight into the phenomenon of interest.
In addition to looking at the macro trade-off produced by varying speed-accuracy emphasis across trial blocks, it is also possible to examine the micro trade-off between speed and accuracy of responding within a particular speedaccuracy emphasis block of the macro function. Models of the macro speed-accuracy trade-off can be differentiated on their predictions regarding the micro trade-off (Pachella, 1974). Osman et al. (2000) presented strong empirical evidence that the macro and micro functions are independent. In their experiment, which used psychophysiological measures as well as behavioral measures, the effect of the macro tradeoff manipulation on RT was independent of that of the micro trade-off, with the micro trade-off affecting the part of the RT interval prior to the lateralized readiness potential (an indicator of readiness to make a left or right response, described later) and the macro trade-off affecting the part of the RT interval after the lateralized readiness potential.
The best models currently for characterizing both RT and accuracy data are sequential sampling models, which assume that information gradually accumulates until a response criterion is reached (Van Zandt, Colonius, & Proctor, 2000). In random walk models, a single counter records evidence as being toward one response criterion and away from another, or vice versa. In race models, separate counters accumulate evidence for each response alternative until the winner reaches criterion. Sequential sampling models explain the speed-accuracy trade-off by assuming that the response criteria are placed further from or closer to the starting point of the accumulation process. They explain biases toward one response over another in terms of asymmetric settings of the response criteria for the respective alternatives. Although continuous models of this general type describe the relation between speed and accuracy well, discrete models that allow pure guesses on a certain percentage of trials can also explain this relation.
Psychophysiological Measures
In recent years, there has been increasing use of psychophysiological measures to supplement RT data (Rugg & Coles, 1995). One of the most popular methods is to record electroencephalograms (EEG), which measure voltage changes in the brain over time from electrodes placed on the scalp. Of particular concern are event-related potentials (ERPs); these are voltage changes in the EEG elicited by a specific event (e.g., a stimulus onset), averaged across many trials to remove background EEG activities. One reason for the popularity of ERPs is that, while a task is being performed, they provide continuous measures of brain activity presumed to be systematically related to cognitive processes. By comparing the effects of task manipulations on various ERP components, their onset latencies, and their scalp distributions, one can make relatively detailed inferences about the cognitive processes. These inferences can be used, along with behavioral measures, to evaluate alternative information processing models.
There are a number of different ERP components, or features, that are indicators of different aspects of processing. These are labeled according to their polarity, positive (P) or negative (N), and their sequence or latency. Early components such as P1 and N1 (the first positive and negative components, respectively) are associated with early perceptual processes. They are called exogenous components because they occur in close temporal proximity to the stimulus event and have a stable latency with respect to it. Later components such as P3 (or P300) reflect cognitive processes such as attention. These components are called endogenous because they are a function of the task demands and have a more variable latency than the exogenous components. For example, when an occasional target stimulus is interspersed in a stream of standards, the P3 is observed in response to targets, but not to standards.
A measure that has been used extensively in studies of action selection is the lateralized readiness potential (LRP; Eimer, 1998), mentioned previously. This potential can be recorded in choice-reaction tasks that require a response with the left or right hand. It is a measure of differential activation of the lateral motor areas of the visual cortex that occurs shortly before and during execution of a response. The asymmetric activation favors the motor area contralateral to the hand making the response, because this is the area that controls the hand. Of importance, the LRP has been obtained in situations in which no overt response is ever executed, allowing it to be used as an index of covert, partial response activation. The LRP is thus a measure of the difference in activity from the two sides of the brain that can be used as an indicator of covert reaction tendencies, to determine whether a response has been prepared even when it is not actually executed. It can also be used to determine whether the effects of a variable are prior or subsequent to response preparation, as Osman et al. (2000) did. Falkenstein, Hohnsbein, and Hoormann (1994) suggested that the latency of the LRP is linked most closely to central decision processes (i.e., action selection), whereas the peak is more closely related to central motor processes.
Electrophysiological measurements and recordings of magnetic fields do not have the spatial resolution needed to provide precise information about the brain structures that producetherecordedactivity.Recentlydevelopedneuroimaging methods, including positron-emission tomography (PET) and functional magnetic resonance imaging (fMRI), measure changes in blood flow associated with neuronal activity in different regions of the brain.These methods have poor temporal resolution but much higher spatial resolution than the electrophysiological methods. Combined use of neuroimaging and electrophysiological methods provides the greatest degree of both spatial and temporal resolution (Mangun, Hopfinger, & Heinze, 1998).
Relevant Stimulus Information
Uncertainty and Number of Alternatives: The Hick-Hyman Law
Merkel (1885), described in Woodworth (1938), provided the initial demonstration that RT increases as a function of the number of possible alternatives. In Merkel’s experiment, the Arabic numerals 1–5 were assigned to the left hand and the Roman numerals I–V to the right hand, in left-to-right order. Results showed that when the number of alternatives increased from 2 to 10 choices, mean RT increased from approximately 300 ms to a little over 600 ms.
Contemporary research dates from Hick’s (1952) and Hyman’s (1953) studies in which the increase in RT with number of alternatives was tied to information theory, which quantifies information in terms of uncertainty (for N equally likely alternatives, the number of bits of information is log2 N). The stimuli in Hick’s study were 10 lamps arranged in an irregular circle, and responses were 10 keys on which the fingers of the two hands were placed. In Hyman’s study, the stimuli were eight lights corresponding to the eight corners of inner and outer squares, and each light was assigned a spoken name. In both studies, RT increased as a logarithmic function of the number of alternatives. Moreover, RT also varied systematically as a function of the relative proportions of the stimulus-response (S-R) alternatives, the sequential dependencies, and speedaccuracy trade-off, as expected on the basis of information theory. This relation between RT and the stimulus information that is transmitted in the responses is known as the Hick-Hyman law:
RT = a + bHT,
where a is basic processing time and b is the amount that RT increases with increases in the amount of information transmitted (HT; log2 N for equally likely S-R pairs with no errors).
The slope of the Hick-Hyman function is negatively correlated with measures of intelligence, which several researchers have claimed to reflect ability to process information rapidly (see Jensen, 1980). However, the fact that the slope of the function is highly dependent on the amount of practice (described later) and other factors severely limits any conclusions that can be drawn from the negative correlation with intelligence tests. A recent study by Vickrey and Neuringer (2000) showed that the Hick-Hyman function has a lower slope for pigeons than for humans, even when they are tested in similar circumstances, which, if the relation to intelligence were accepted, would imply that pigeons are more intelligent than humans.
One criticism of the Hick-Hyman law is that the function relating RT to number of alternatives is not logarithmic. Kvälseth (1980) introduced a variety of laws, including a power law for the case of equally likely alternatives and an exponential law for cases in which the alternatives are not equally probable. Longstreth, El-Zahhar, and Alcorn (1985) claimed that the specific power law, RT = a + b(1 – N–1), provides a better fit to data for equiprobable alternatives than the logarithmic function. Longstreth et al.’s main argument for the power law is that as the number alternatives increases beyond 8, the function is no longer linear with respect to the logarithm, but becomes curvilinear (see Longstreth, 1988). Although theoretically derived from an attentional model, Longstreth et al.’s power law is a special case of the more general power law proposed by Kvälseth (1980). In addition, Kvälseth (1989) and Welford (1987) pointed out that Longstreth et al.’s power law has several problems. Kvälseth (1989) captures the status of the Hick-Hyman law, stating, “Although, on purely empirical grounds, Hick-Hyman’s law may not be uniformly superior to other lawful relationships, it has been clearly established that it does provide a good summary description of a substantial amount of data” (p. 358).
Stimulus-Response Compatibility
Stimulus-response compatibility (SRC) is one of the principal factors affecting efficiency of action selection. SRC refers to the fact that performance is better with some mappings of stimuli to responses than with others. SRC effects are ubiquitous and occur with a variety of stimulus and response sets, although much of the research has focused on spatial SRC effects.
Spatial Compatibility Effects
Paul Fitts is given credit for formalizing the concept of SRC. Fitts and Seeger (1953) examined performance of eight-choice tasks using all combinations of three stimulus arrangements and three response arrangements. They found that responses were faster and more accurate when the stimulus and response arrangements corresponded spatially than when they did not. Fitts and Deininger (1954) showed that for conditions in which the stimulus and response arrangements were the same, responses were much slower with an arbitrary mapping of S-R locations than with one in which the corresponding response was made to each stimulus. Even more interesting, performance was also much better with a mirrorreverse mapping of stimulus locations to response locations than with a random mapping, although performance was still inferior to that of the spatially corresponding mapping.
The spatial SRC effect is robust in that it is obtained with auditory and tactual stimuli and with key presses, joystick movements, and unimanual aimed movements (see Proctor & Reeve, 1990, and Hommel & Prinz, 1997, for edited volumes on SRC). The slope of the function for the Hick-Hyman law, relating RT to the number of alternatives, is inversely related to SRC (Smith, 1968), approaching zero for highly compatible S-R mappings (Teichner & Krebs, 1974). In other words, SRC effects increase in magnitude as the number of S-R alternatives increases.
Many studies have used a two-choice task in which a left or right key press is made to a left or right stimulus. In twochoice tasks, responses are typically 50–100 ms faster when the S-R mapping is spatially compatible than when it is not, regardless of whether the stimuli are visual or auditory. Moreover, PET scans show increased bloodflow for incompatible mappings compared to compatible mappings in the same brain regions (left rostral dorsal premotor and posterior parietal areas) for both visual and auditory modalities (Iacoboni, Woods, & Mazziotta, 1998). This spatial SRC effect is a function of relative position of the stimuli and responses: It occurs even when the stimulus display or hands are shifted to the left or right of center (Nicoletti, Anzola, Luppino, Rizzolatti, & Umiltà, 1982). Moreover, the SRC effect is found when the hands are crossed so that the left hand operates the right key and the right hand the left key (Roswarski & Proctor, 2000), as well as when the responses are made with two fingers on the same hand (Heister, Schroeder-Heister, & Ehrenstein, 1990). The dependence of the effect on the spatial relations of the stimuli and responses has led most accounts of spatial SRC to focus on spatial coding as its basis. The spatial codes are based on the task goals, as illustrated in a study by Riggio, Gawryszewski, and Umiltà (1986) in which subjects operated the left key with a stick held in the right hand and the right key with a stick held in the left hand. Even though the hands were on their normal sides, responses were faster with the S-R mapping in which the stimuli corresponded to the location of the response key and not the hand used for responding.
Conceptual, Perceptual, and Structural Similarity
A variety of SRC effects in addition to spatial compatibility have been demonstrated. Kornblum, Hasbroucq, and Osman (1990) and Kornblum and Lee (1995) have argued that SRC effects will occur for any situation in which the stimulus and response sets have dimensional overlap (i.e., are similar). Dimensional overlap is presumed to include both conceptual and perceptual similarity. The role of conceptual similarity is illustrated in the findings that spatial SRC effects, broadly defined, occur when location words are spoken in response to physical location stimuli, as well as when left-right key presses are made to the words left and right or to left- and right-pointing arrows. The role of perceptual similarity is shown by the finding that SRC effects are larger within the spatial-manual and verbal-vocal modes, that is, for physical locations mapped to key presses and location words mapped to naming responses, than between the modes (Wang & Proctor, 1996).
SRC effects are also obtained when the S-R sets do not share conceptual or perceptual similarity but have structural similarity. When an ordered set of stimuli (e.g., A, B, C, D) is mapped to an ordered set of responses (e.g., 1, 2, 3, 4), RT is shorter for a mapping that preserves or reverses this order than for one that does not. Another type of structural compatibility effect occurs when a symbolic two-dimensional stimulus set is mapped to index and middle finger responses on each hand. When two letters (O, Z) of two sizes (large or small) are mapped to the responses, the left-to-right mapping of O, o, z, Z is easier than one of O, z, o, Z (Miller, 1982; Proctor & Reeve, 1985). Proctor and Reeve presented evidence that this difference is due to the letter identity distinctions being salient for the stimulus set and the distinctions between the two left and two right responses being salient for the response set. Performance is best for the condition in which the salient stimulus feature maps directly onto the salient response feature. In other words, translation of the specific stimulus into a response can occur more quickly when salient features correspond. Salient features coding has been shown to determine the compatibility effects obtained for a variety of situations in which the stimulus and response sets have structural similarity, but no conceptual or perceptual similarity (Proctor & Reeve, 1990).
Compatibility Effects in Two Dimensions
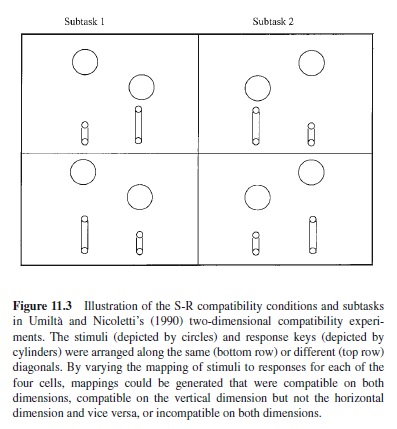
Umiltà and Nicoletti (1990) examined compatibility along two dimensions in a two-choice task by varying the stimulus and response locations for a set of trials along a diagonal (see Figure 11.3). They found that the compatibility effect was larger for the horizontal dimension than for the vertical dimension, a phenomenon they called right-left prevalence. Vu and Proctor (2001) showed that this right-left prevalence effect can be reversed to top-bottom prevalence by increasing the relative salience of the vertical dimension. This was accomplished by using response sets that emphasized the top-bottom distinction. In one experiment that showed top-bottom prevalence, subjects responded with anatomical top-bottom effectors, a hand and foot. In another experiment, top-bottom prevalence was obtained when one hand was placed over the other so that the top-bottom distinction was salient. Thus, although right-left prevalence typically is obtained when left-right effectors are used, and top-bottom prevalence when top-bottom effectors are used, the prevalence effects do not seem to have an anatomical basis, but are by-products of the relative salience of the two dimensions.
Compatibility effects can occur as well when the spatial dimension along which the stimulus locations vary is orthogonal to that along which the response alternatives vary. For top-bottom stimuli mapped to left-right key press or vocal responses, the mapping of top-right and bottom-left yields faster responding than the alternative mapping (Cho & Proctor, 2001). A variant of salient features coding can also explain this mapping effect (Weeks & Proctor, 1990). Specifically, evidence indicates that the two alternatives on the vertical and horizontal dimensions are coded asymmetrically, with top and right being the polar referents for their respective dimensions. Consequently, the salient features coding explanation is that action selection occurs faster for the top-right/bottom-left mapping than for the alternative mapping because it is the one for which the salient features correspond. Adam, Boon, Paas, and Umiltà (1998) proposed that this asymmetric coding is a property of verbal codes but not spatial codes. However, Cho and Proctor provided evidence that it is a general property of spatial coding.
With unimanual movements of a joystick or finger, the top-right/bottom-left mapping is also typically more compatible than the alternative mapping. In this case, though, the mapping preference is affected by the location of the response apparatus. The top-right/bottom-left advantage is enhanced when responding in the right hemispace, but it reverses to a top-left/bottom-right advantage when responding in the left hemispace (Weeks, Proctor, & Beyak, 1995). Lippa (1996) provided evidence that the mapping preference is also affected by hand posture. According to her referential coding hypothesis, the finger-to-wrist axis provides a reference frame that allows the response set to be coded parallel to the stimulus set. For example, when left-right responses are made with the right hand held at a comfortable 45–90º, the left response can be coded as top and right response as bottom. Referential coding can explain many results obtained with unimanual responses, but it cannot explain why the mapping pBibliography: described above occur when the hand and finger are in a neutral posture that allows only leftright deflections perpendicular to the sagittal body midline (Michaels & Schilder, 1991).
Because of this deficiency of the referential coding hypothesis, Lippa andAdam (2001) proposed an end-state comfort hypothesis. Similar to referential coding, the end-state comfort hypothesis views orthogonal compatibility as a correspondence effect. However, it assumes that the response dimension is mentally rotated, according to relative hand posture, to bring it into alignment with the stimulus dimension.
The direction of rotation, clockwise or counterclockwise, is determined by physical constraints of the body. The response dimension is mentally rotated in the direction that would yield the most comfortable end-state posture if the hand were actually rotated (inward movement for the left or right hand when positioned at centered or ipsilateral locations, and outward movement when positioned at contralateral locations). The end-state comfort hypothesis can account for more results obtained with unimanual responses than the referential coding hypothesis, but both hypotheses are not directly applicable to the orthogonal compatibility effects obtained with bimanual or vocal response sets.
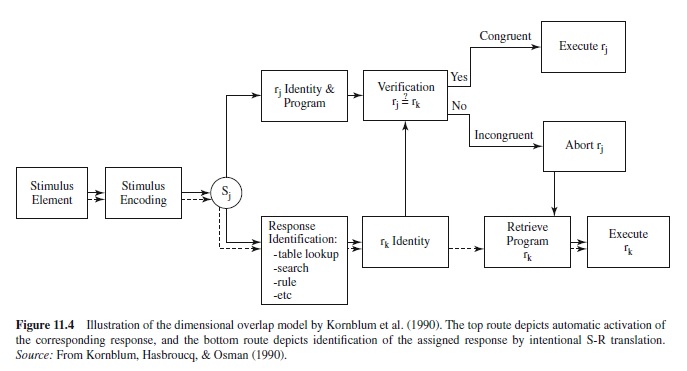
Dual-Route Models
Virtually all explanations of SRC effects agree that at least part of the difference in RT between compatible and incompatible mappings involves the time to translate the stimulus into its assigned response based on the instructions provided for the task. Translation is presumed to be fastest when an identity rule can be applied (i.e., make the response corresponding to the stimulus), intermediate when some other rule can be used (e.g., make the response that is the mirror opposite of the stimulus), and slowest when the response must be retrieved via the specific S-R associations defined for the task. Although some models rely exclusively on intentional translation (e.g., Rosenbloom & Newell, 1987), dual-route models that propose an additional direct (or automatic) response-selection route have come to be favored (e.g., Kornblum et al., 1990; see Figure 11.4). The basic idea is that when a stimulus occurs it tends to produce activation of its corresponding response by way of long-term S-R associations, regardless of the S-R mapping defined for the task. The resulting activation produces a benefit in responding when the corresponding response is correct, but a cost when it is not. The major reason that dual-route models have become popular is that correspondence effects often occur for irrelevant stimulus dimensions (see Lu & Proctor, 1995), as discussed in a subsequent section.
Sequential Effects
Repetition Benefit
Bertelson (1961) was the first to formally investigate sequential effects on performance. He showed that for a two-choice task, in which left-right stimuli were mapped compatibly to left-right keys, the total response time for a set of trials was less when the proportion of repetitions was .75 than when it was .25. This repetition benefit was evident when the response-stimulus interval (RSI) was 50 ms but not when it was 500 ms.
Since Bertelson’s (1961) study, numerous, more detailed investigations of sequential effects in choice-reaction tasks have been conducted. First-order sequential effects are those that involve the relation of the current trial to the immediately preceding trial. The most common first-order effect is that the response to a stimulus is faster when the S-R pair for a trial is a repetition of the preceding S-R pair than when it is not. In two-choice tasks, this repetition benefit is obtained only when the RSI is short. At RSIs of 500 ms or longer, a benefit for alternations over repetitions is typically found instead. The repetition benefit is larger in tasks with more than two choices, being an increasing function of the number of S-R alternatives, and in these tasks a repetition benefit is found even at long RSIs (Soetens, 1998). The first-order sequential effects have been attributed to two processes, much like those proposed for priming effects (Neely, 1977). At short RSIs, residual activation from the preceding trial produces automatic facilitation when the current trial is identical to it; at long RSIs, strategic expectancy regarding the nature of the next trial produces faster responses for expected than unexpected stimuli (Soetens, 1998). This expectancy is for the alternative S-R pair in two-choice tasks, but for repetition of the same pair in tasks with more alternatives.
Pashler and Baylis (1991) evaluated the locus of the repetition benefit for tasks in which two stimuli were assigned to left, middle, and right response keys operated by index, middle, and ring fingers of the right hand. Two of the stimuli were digits, two were letters, and two were nonalphanumeric symbols (e.g., & and #). Stimuli were mapped to responses in a categorizable (e.g., digits-to-left response, letters-tomiddle response, and symbols-to-right response) or uncategorizable (e.g., a digit and a letter to the left response, etc.) manner. For both mappings, the repetition benefit occurred primarily when the same stimulus was repeated and not when only the response was repeated. This repetition benefit for the same stimulus was not found when responses on alternate trials were vocal and manual. Consequently, Pashler and Baylis concluded that the repetition effects were at the stage of response selection, with the normal responseselection process being bypassed when the stimulus and response were repeated.
In Pashler and Baylis’s (1991) experiments, a benefit for response repetition alone tended to occur with categorizable but not uncategorizable S-R mappings. Campbell and Proctor (1993) verified this effect, showing a benefit of approximately 40 ms for response repetition alone with categorizable mappings but not uncategorizable mappings. Their remaining experiments showed that this response repetition benefit, as well as the additional benefit for repeating the same stimulus, could be obtained when the responses on successive trials were made with different hands. In the critical conditions, the stimuli were presented to the left or right of fixation on alternate trials, with responses to the left stimulus made with the three fingers on the left hand and responses to the right stimulus made with the three fingers on the right hand. A cross-hand repetition benefit was obtained when either spatial or finger information was consistent across hands, but not when both consistencies were eliminated. These results imply that the response sets can be coded in terms of locations or effectors and that response selection benefits from repetition of the stimulus category when it maps onto a salient feature of the response sets.
Soetens (1998) examined sequential effects for tasks in which subjects responded to four stimuli located at the corners of an imaginary square by pressing the left key if the stimulus was to one side and the right key if it was to the other. When left-right stimulus locations were mapped compatibly to left-right responses, the repetition benefit at the short RSI (50 ms) was primarily associated with the response (i.e., the benefit was evident when the stimulus side was the same as on the previous trial, but the location was different). At the long RSI (1,000 ms), a small alternation benefit was evident. With an incompatible S-R mapping (i.e., left side to right response), the results were similar, but with an increased benefit for repeating the same stimulus, particularly at the short RSI. When up-down responses were made to the left-right stimulus locations, response and stimulus repetition benefits of similar magnitudes were found at the short RSI. At the long RSI, the only effect was a repetition benefit for the same stimulus. Soetens concluded that automatic facilitation shifted toward stimulus-related processes as the mapping became less compatible. Together, the studies of Pashler and Baylis (1991), Campbell and Proctor (1993), and Soetens indicate that response repetition, without stimulus repetition, is beneficial when there is a structural relation between the stimulus and response sets and that repetition of the stimulus is more important when the mapping is arbitrary.
Although first-order sequential effects have been most widely studied, second- and third-order repetition effects, involving the sequence of the preceding two or three stimuli, respectively, are larger and more consistent (Soetens, 1998). For two-choice tasks, at short RSIs, RT benefits from multiple repetitions, regardless of whether the present trial is a repetition or an alternation. For example, responses on the current trial tend to be faster if the three preceding trials were repetitions than if they were alternations. At long RSIs, however, a prior string of repetition trials is beneficial if the current trial is also a repetition, but a prior string of alternation trials is beneficial if the current trial is an alternation. These two patterns of results can be attributed to automatic activation and subjective expectancies, respectively. The higher order effects in Soeten’s study also showed the patterns indicative of automatic facilitation at the short RSI and subjective expectancy at the long RSI.
Is the Hick-Hyman Law an Artifact of Repetition Effects?
Kornblum (1967, 1968) noted that, unless explicitly controlled, the proportion of repetition trials decreases as set size increases. Therefore, he proposed that the Hick-Hyman law is an artifact of repetition effects. Kornblum (1968) used a four-choice task in which four lights were mapped to four response keys and information was varied by manipulating stimulus probabilities. For three levels of information, conditions were constructed in which the probability of repetition was high or low. RT was shorter for the high-repetition conditions than for the corresponding low-repetition conditions, and these latter conditions showed only a nonsignificant effect of information on RT. Kornblum (1967) conducted a similar experiment in which the number of alternatives was two, four, or eight. For four- and eight-choice tasks, RT was shorter on repetition than on nonrepetition trials, with the slope being less for repetition trials. Within these tasks, RT for repetition trials increased as the amount of stimulus information increased, but RT for nonrepetitions did not.
Hyman and Umiltà (1969) noted that the RSI in Kornblum’s (1967, 1968) experiments was approximately 140 ms, a short interval that would maximize repetition effects and minimize preparation for the subsequent trial. They replicated three of Kornblum’s (1968) conditions, but used an average RSI of 7.5 s. Although RT was faster for repetition than nonrepetition trials, the slopes of the two functions were approximately equal. Hyman and Umiltà concluded, “There seems little doubt that the information hypothesis is much more compatible with our results than those of Kornblum’s” (p. 47). In other words, the Hick-Hyman function is not an artifact of the proportion of repetition trials when there is adequate preparation time.
Advance Information
Warning Effects
Preparation is usually studied by presenting a neutral warning signal at various intervals prior to the onset of the imperative stimulus. Bertelson (1967) had subjects press a right key to a right light and a left key to a left light. The warning signal was an auditory click that, in different blocks, occurred 0, 20, 50, 100, 150, 200, or 300 ms prior to the visual stimulus. At the 0-ms warning interval, RT was approximately 265 ms. It decreased to a minimum of 245 ms at the 150-ms interval and then increased slightly to 250 ms at the two longest intervals. However, the error rate increased from about 7% at the shorter intervals to 12% at the 100- and 150-ms intervals, and decreased slightly to 9% at the longer intervals. Thus, the effect of the warning signal was to increase readiness to respond quickly, but at the expense of accuracy.
Posner, Klein, Summers, and Buggie (1973) obtained similar results for a two-choice task in which the compatibility of the mapping of the stimulus locations to responses was manipulated. Each trial was preceded by no warning or a 50-ms warning tone, followed at intervals of 50, 100, 200, 400, and 800 ms by a stimulus to the left or right of fixation. RT was a U-shaped function of foreperiod, reaching a minimum at the 200-ms interval. Error rate showed an opposing, inverted U-shaped function, being highest at the 100-ms interval. The main effect of compatibility was significant in the RT and error data, but compatibility did not interact with foreperiod. These results suggest that the warning tone altered alertness, or readiness to respond, but did not affect the rate at which the information built up in the response-selection system.
RT continues to increase as the foreperiod increases beyond 800 ms, up to at least 5 s. Sanders and Wertheim (1973) failed to find an effect of foreperiod between 1 and 5 s for auditory stimuli, although they found the standard increase in RT for visual stimuli. However, Sanders (1975) demonstrated that the critical factor seems to be stimulus intensity: Auditory stimuli below 70 dB showed foreperiod effects similar to those shown by visual stimuli, and there was a trend toward smaller effects for high-intensity visual signals.
Precuing Effects
Leonard (1958) was the first to demonstrate that subjects can use advance information to prepare for a subset of S-R alternatives. He tested himself in a six-choice reaction task in which six stimulus lights were mapped compatibly to six response keys pressed by the fingers of each hand. In the sixchoice condition, all six stimuli were lit, and the target light went off 100 ms later. In a three-choice condition, only the left or right set of three stimuli was used. Of most interest was a precue condition in which the subject did not know whether the choice would involve the three left locations or the three right locations until the lights designating those locations were lit (i.e., those locations were precued). RT decreased as a function of the precuing interval, with RT at the 500-ms interval being equivalent to that of the three-choice task.
Subsequent studies using four-choice tasks have obtained similar results, in which the benefit for precuing the two left or two right locations occurs within the first 500 ms of precue onset (Miller, 1982; Reeve & Proctor, 1984). However, when other pairs such as alternate locations are precued, the maximal benefit is not evident until a longer interval. Reeve and Proctor (1984) showed that the advantage for precuing the two left or two right locations does not depend on the fact that they typically involve responses from different hands.With an overlapped hand placement in which the index and middle fingers from the two hands are alternated, the two left or right locations show a similar precuing advantage relative to other pairs of locations. These and other findings imply that the time needed to obtain the maximal benefit from a precue varies as a function of how long it takes to translate the precue information. Proctor and Reeve (1986) attributed this pattern of differential precuing benefits to the salience of the left-right distinction.
Kantowitz and Sanders (1972) distinguished between two types of precue: utility and necessity. Utility precues, as in the studies just discussed, are helpful in reducing the number of alternatives, but do not provide information that is necessary for responding. Necessity precues tell subjects what information is relevant for the current trial (e.g., whether they are to respond to stimulus color or shape). RT is longer when the precue is a necessity than when it is only useful. Because the information provided by necessity precues must be used at all intervals, it is more difficult to respond at shorter ISIs. With utility precues, subjects use the information at longer intervals but not shorter ones.
Relevant and Irrelevant Stimulus Information
Noncorrespondence of Relevant and Irrelevant Information
Effects of irrelevant information on performance have been studied extensively in many areas of experimental psychology. Three such effects studied in the choice reaction literature—the Stroop color-naming effect, the Eriksen flanker effect, and the Simon effect—involve correspondence of relevant and irrelevant stimulus information.
The Stroop Effect
The best-known example of irrelevant information affecting response selection is the Stroop color-naming task (see MacLeod, 1991, for a review). In this task, color words are presented in different ink colors, and subjects are instructed to name the ink color while ignoring the color word. In Stroop’s (1935/1992) study, subjects took 110 s to name a list of 100 colors presented in incongruent color words, compared to 63 s to name a list of 100 colors presented in solid squares. Thus, conflicting color words nearly doubled the naming time, a phenomenon known as the Stroop effect. Stroop also reported that the time to read 100 color words in incongruent ink colors was 43 s, compared to 41 s when the words were presented in black ink. Thus, the interference with color naming was asymmetric: Irrelevant words interfered with naming ink colors, but irrelevant ink colors did not interfere with reading color words.
This asymmetric pattern of interference has been reported in numerous subsequent studies, including versions of the task in which RTs to individual stimuli are recorded. An important finding is that the pattern of asymmetry is dependent on the response mode. When the task involves pointing to a matching color, responses to color words are delayed by incongruent colors, but responses to colors are not delayed by irrelevant color words (Durgin, 2000). Similarly, in spatial versions of the task, in which the word left or right is presented in left or right locations or with an arrow pointing to the left or right, the words produce interference when the responses are made vocally, but the locations or arrows produce interference when the responses are key presses (Lu & Proctor, 1995).
Stroop (1935/1992) showed in his Experiment 3 that a dimension that does not produce interference (e.g., ink colors when the task is word reading) can be made to do so with practice. In his experiment, subjects practiced four lists of 50 words in the color-naming task for 8 days. The average time to read the list decreased from 50 s on the first day to 33 s on the last day, but this was still longer than the 25 s to name a neutral list of colored swastikas. Subjects also performed the word-reading task prior and subsequent to practicing the color-naming task. The time to perform the word-reading task was nearly twice as long (35 s) after the color-naming practice as before (19 s). Thus, the practice increased the strengths of the associations between colors and names, and the colors now produced interference with reading color words.
More generally, relative strength of association is a good predictor of whether an irrelevant stimulus dimension will affect responding to a relevant stimulus dimension. Lu and Proctor (2001) classified the association of stimulus dimensions to key presses as high if they were both conceptually and perceptually similar (e.g., arrows are spatial and nonverbal, as are key presses), intermediate if they were only conceptually similar (e.g., location words are spatial but verbal), and low if they were neither (e.g., colors and color words are not similar to key presses). Across several experiments using various combinations of relevant and irrelevant stimulus dimensions, the relative magnitudes of effect size were predictable based on relative association strength. Baldo, Shimamura, and Prinzmetal (1998) obtained similar results varying response modalities in addition to stimulus dimensions: Robust Stroop effects to location word/arrow stimuli were observed when responding manually to location words or vocally to arrows, but not for the reverse relations. The results of Lu and Proctor and of Baldo et al. are generally consistent with Kornblum et al.’s (1990) emphasis on response activation varying as a function of dimensional overlap and with parallel distributed processing models of the type proposed by Cohen, Dunbar, and McClelland (1990), which rely on relative association strength.
The Eriksen Flanker Effect
Another widely studied effect of irrelevant information is the Eriksen flanker effect (Eriksen & Eriksen, 1974). In the typical experiment examining this effect, one or more stimuli are assigned to left-right responses. The target letter for each trial is presented at a known, centered location and is flanked by instances of a distractor letter. In Eriksen and Eriksen’s experiment, the letters H and K were assigned to one response and the letters S and C to the other response. The flanking letters could be the same as the target (HHHHHHH), the letter assigned to the same responses as the target (congruent; KKKHKKK), or a letter assigned to the opposite response (incongruent; SSSHSSS or CCCHCCC). When the letters were in close spatial proximity, responses were faster when the flanking letters were identical to or congruent with the target than when they were incongruent. This congruency effect decreased as the spatial separation between the letters increased.
Because distractors that are not potential targets produce little or no interference, the results suggest that the effects reflect response activation. That is, the flanking letters activate the response to which they are assigned, producing response competition when that response is not the one signaled by the target. This competition is evident in a tendency for the lateralized readiness potential to show initial activation of the wrong response 150 to 250 ms after onset of the target and incongruent distractors (Gratton, Coles, Sirevaag, Eriksen, & Donchin, 1988). Eriksen and Schultz (1979) proposed a continuous flow account of the flanker effect, much like McClelland’s cascade model, in which stimulus information gradually accumulates in the visual system and continuously flows into the response system. Initially, a wide range of responses is activated, but as the output from the perceptual system becomes more exact, the response activation becomes increasingly restricted to the appropriate response. This account assumes that after a flanking letter is fully identified, it will no longer produce response activation. However, if it is assumed that fully identified flankers may still contribute to response activation, then discrete stage models can account for the results as well (Mordkoff, 1996).
The Simon Effect
The Simon effect is another close relative of the Stroop effect (Lu & Proctor, 1995). In the typical Simon task, stimulus location is irrelevant and the responses, most often left-right key presses, vary along a location dimension. The relevant stimulus dimension typically involves a distinction other than location (e.g., color or letter identity). The Simon effect is that responses are faster when the location of the stimulus and response correspond than when they do not. The effect typically is larger when responses are fast than when they are slow, implying that activation of the location information occurs quickly and then decreases because it is irrelevant to the task (Hommel, 1993b). Consistent with this view, when the correct response is not the one that corresponds with the location of the stimulus, the lateralized readiness potential shows evidence of slight, initial activation of the spatially corresponding response, which then shifts to activation of the correct, noncorresponding response (De Jong, Liang, & Lauber, 1994).
Considerable research on the Simon effect has focused on why stimulus location is coded when it is irrelevant to the task. Stoffer and Umiltà (1997) attribute the Simon effect to shifts of attention associated with eye movements. According to them, the position of the object attended at stimulus onset, typically a fixation point, provides a frame of reference. The location of the stimulus relative to the focus of attention is coded only when attention is shifted to the stimulus. This code specifies the direction and amplitude of the saccade program to shift fixation to the stimulus. The types of evidence they have presented in support of the attention-shifting hypothesis are that the Simon effect is absent when attention shifts are prevented by the need to report a stimulus presented at fixation and reversed when an attention shift back from the stimulus location to the fixation point is required.
Hommel (1993b) has argued instead that spatial coding occurs with respect to various frames of reference, of which the focus of attention may be one. Perhaps the best evidence for his referential coding hypothesis is that the Simon effect can vary as a function of multiple frames of reference. In a procedure used by Lamberts, Tavernier, and D’Ydewalle (1992) and Roswarski and Proctor (1996), a stimulus can occur in one of eight locations, four to the left of fixation and four to the right. Initially, four boxes appear to one or the other side to designate the possible locations for that trial. Then the two left or two right boxes disappear, and the imperative stimulus is presented in one of the remaining boxes. In this case, a Simon effect occurs with respect to three frames of reference: Left-right side of fixation; two left versus two right on a side; and the left-right location within the final pair. The largest difference between corresponding and noncorresponding responses occurs when the stimulus is in the far left or far right location, for which all three spatial codes are in agreement (e.g., all left or all right).
As with compatibility for relevant stimulus information, the Simon effect varies as a function of task goals. Hommel (1993a) had subjects respond to a high or low pitch tone, presented to the left or right side, by pressing a left or right key. The key closed a circuit that lit a light on the opposite side. When instructed to press the left key to the high pitch tone and the right key to the low pitch tone, a typical Simon effect occurred. However, when instructed to turn on the right light to the high pitch tone and the left light to the low pitch tone, the Simon effect was a function of light location. That is, in this case, responses were faster when the stimulus was on the side opposite the responding hand, rather than on the same side. Guiard (1983) obtained a similar finding in an experiment in which subjects responded to tone pitch by turning a steering wheel clockwise or counterclockwise. In the condition of most interest, the subject’s hands were placed at the bottom of the wheel, and a clockwise turn moved a cursor to a right target location and a counter-clockwise turn moved it to the left. Because of the hand placement, when the wheel was turned clockwise the hands moved to the left, and vice versa when the wheel was turned counter-clockwise. A Simon effect was obtained as a function of the direction of wheel rotation, rather than as a function of the direction in which the hands moved.
Another goal-related phenomenon is the Hedge and Marsh (1975) reversal, in which the Simon effect reverses to favor noncorresponding locations when the response keys are labeled according to the same dimension as the relevant stimulus information, and subjects are instructed to respond in an incompatible manner (e.g., press the green key to the red stimulus and vice versa). The explanation proposed by Hedge and Marsh, and which has continued to be the most widely accepted, is that of logical recoding. The basic idea is that a respond opposite rule is applied both to the relevant stimulus dimension and, inadvertently, to the irrelevant location dimension, leading to activation of the noncorresponding response.
Negative Priming
For the Stroop color-naming task, and related tasks with irrelevant stimulus information, the target stimulus value on a trial can not only be a repetition or nonrepetition of the relevant value on the previous trial, but also the same as the value of the irrelevant information. When the value of the relevant stimulus dimension is the same as that of the irrelevant dimension on the preceding trial, an effect called negative priming is often observed. This effect was first demonstrated by Dalrymple-Alford and Budayr (1966) for the Stroop color-naming task. Subjects had to name the ink colors for lists of Stroop color words that differed in the relation between successive stimuli. For the control list, there was no relation between the word or ink color for successive stimuli. For the ignored repetition list, however, the irrelevant color word for one stimulus was the relevant color for the next stimulus. In other words, if the color word for stimulus n-1 was red, then the ink color for stimulus n was red. The finding of interest was that the time to name the colors for the ignored repetition list was much longer than that for the control list. This slowing of responses when the to-be-ignored information on the previous trial is relevant on the current trial is the phenomenon of negative priming.
Negative priming has subsequently been studied most often using a method in which responses to individual stimuli are measured. In that situation, the trials are often presented as pairs, with the first trial called a prime and the second a probe. Negative priming is shown when responses are slower for trials in which the previously irrelevant information is now relevant than for neutral trials. The negative priming effect has been found in a variety of tasks for which irrelevant information is present (Fox, 1995; May, Kane, & Hasher, 1995), including not only tasks that require identification of an object but also those that require localization.
The most straightforward interpretation of negative priming effects is that of selective inhibition: The irrelevant information must be inhibited in order to respond to the relevant information, and this inhibition carries forward to the next trial. Consequently, the response will be slowed if the inhibited information is now relevant. Although numerous findings are consistent with the selective inhibition hypothesis, they can also be accounted for without assuming inhibition. Moreover, the situation has been shown to be much more complex than the selective inhibition hypothesis suggests, and alternative explanations have been proposed. The two most prominent alternatives are feature mismatching and episodic retrieval. According to the feature mismatch hypothesis (Park & Kanwisher, 1994), symbol identities are bound to objects and locations, and any change in the bindings from the previous trial will produce negative priming. The episodic retrieval hypothesis (Neill & Valdes, 1992) states that presentation of a stimulus evokes retrieval of previous episodes involving the stimulus. Because recent episodes are most likely to be retrieved, if the target stimulus was a distractor on the previous trial, the episode retrieved will include an ignore tag.
One problematic finding for the inhibition account is that negative priming effects do not appear to be short-lived. DeSchepper and Treisman (1996) found negative priming after a delay of 30 days between the prime and probe trials. In addition, negative priming depends on the relation between the prime and probe trials. For example, for the Stroop task, the effect is not found if the probe stimulus that follows the prime Stroop stimulus is a color patch and not a colored word (Lowe, 1979). A simple inhibition account would seem to predict negative priming in this situation as well as in that for which the probe stimulus was a colored word.
Multiple Tasks
Task Switching
In his classic monograph, “Mental Set and Shift,” Jersild (1927) began by saying, “The fact of mental set is primary in all conscious activity. The same stimulus may evoke any one of a large number of responses depending upon the contextual setting in which it is placed” (p. 5). Jersild conducted experiments in which subjects made a series of judgments regarding each stimulus in a list as a function of whether a single task was performed for all stimuli or two tasks were performed in alternating order. The major finding was that in many situations the time to complete the list was longer for mixed lists than for pure lists of a single task.
Beginning in the mid-1990s, there has been a resurgence of interest in task switching. Research conducted on task switching, in which two tasks are presented in a fixed order (e.g., on alternate trials), has suggested that there are two components associated with changing the task set from the previous trial. One component involves voluntary preparation for the forthcoming trial, with responses for the next trial becoming progressively faster as the RSI increases. However, time to prepare for the new task cannot be the only factor contributing to the switching cost, because the cost is still evident when the RSI is long (Allport, Styles, & Hsieh, 1994; Rogers & Monsell, 1995). A second component, which Allport et al. (1994) called task set inertia and Rogers and Monsell (1995) called exogenous task set reconfiguration, is not under the subject’s control. Apparently only a single trial with the new task is necessary to complete configuration for that task. Rogers and Monsell (Experiment 6) used sequences of four task repetitions and then a switch to the alternate task for four consecutive trials, and so on, and found that the switch costs were eliminated after the first trial of the new task.
Shaffer (1965) conducted a study in which trials with compatible and incompatible spatial mappings were randomly mixed. The stimulus to which the subject was to respond occurred in a left or right location, and a centered horizontal or vertical line signaled whether the mapping for the trial was compatible or incompatible. When the mapping signal occurred simultaneously with the stimulus, the standard spatial compatibility effect was eliminated. Vu and Proctor (2001) used stimulus color to designate the mapping and obtained similar results with left-right physical-location stimuli, as Shaffer used, as well as with left-right pointing arrows. These findings are consistent with the fact that, in a variety of situations, performance of the easier of two tasks is harmed more by mixing (Los, 1996). However, Vu and Proctor found that when the stimuli were the words left and right, the advantage for the compatible mapping was enhanced compared to pure blocks of one trial type. These results, along with many others, suggest that words are processed differently than physical locations and arrows.
Proctor and Vu (2002) also showed that mixing locationrelevant and location-irrelevant trials within a trial block alters the stimulus-response compatibility (SRC) effects obtained for each task. When physical location stimuli were used to convey the location information, the standard SRC effect was eliminated for location-relevant trials. However, the SRC effect was not affected with arrow stimuli and was enhanced with location word stimuli. Mixing the two trial types also affects the Simon effect obtained for the locationirrelevant trials. For all stimulus types, when the locationrelevant mapping was compatible, the Simon effect was enhanced compared to pure blocks of Simon trials; when the location-relevant mapping was incompatible, a reverse Simon effect was obtained. With arrows and words, the reverse effect was smaller than the positive effect. However, with physical locations, the reverse Simon effect was at least as large as the positive effect obtained with the compatible location-relevant mapping. This outcome implies that there was no automatic activation of the corresponding response. The reversal for physical location stimuli obtained when the location-relevant mapping was incompatible was evident even when the trial type was precued by up to 2.4 s before presentation of the stimulus. This outcome indicates that the reversal does not reflect only a strategy of preparing the noncorresponding response in anticipation that location may be relevant to the trial.
Psychological Refractory Period
In a common dual-task procedure, subjects perform two different choice-reaction tasks, Task 1 (T1) and Task 2 (T2), on a single trial. The stimulus onset asynchrony (SOA) between the stimuli for T1 (S1) and T2 (S2) is varied. The typical finding is that RT for the second task (RT2) is slowed as the SOA decreases. Telford (1931) called this phenomenon the psychological refractory period (PRP) effect. Extensive research on the PRP effect has been conducted over the past 50 years, and explanations have been proposed in terms of informationprocessing bottlenecks, demands on limited capacity resources, and strategies adopted to satisfy task constraints (Meyer & Kieras, 1997; Pashler, 1998). The most widely accepted account in recent years is a response-selection bottleneck model advocated by Pashler and colleagues (see Figure 11.5). According to this model, stimulus identification and response execution occur in parallel for the two tasks. However, response selection operates serially because it requires a single-channel mechanism.

The evidence for the response-selection bottleneck model comes primarily from using locus of slack logic (Schweickert, 1983) to interpret the patterns of additive and interactive effects produced by variables presumed to selectively affect stimulus identification, response selection, and response execution. According to the model, identification of S2 commences immediately upon its presentation, regardless of the SOA. At long SOAs response selection can begin as soon as stimulus identification is completed, but at short SOAs it cannot begin until response selection for T1 is finished. Consequently, there is slack in the processing sequence for T2 between the completion of stimulus identification and initiation of response selection. At short SOAs, the slack can absorb, at least in part, an increase in time to identify S2. This leads to the effect of the stimulus-difficulty manipulation being smaller at the short SOAs than at the long SOAs. In contrast, for variables that affect response selection or response execution, which have their influence after the bottleneck, the extra time cannot be absorbed by the slack, and, therefore, their effects should be additive with those of SOA. These predicted patterns of results have been found for several variables of the respective types.
Meyer and Kieras (1997) have mounted a challenge to the response-selection bottleneck model, arguing that evidence supporting it reflects a strategy adopted by subjects when the instructions state or imply that the response for T1 must be made before that for T2. They propose that there is no capacity limitation in processing other than a bottleneck for response execution when the tasks require responses from the same output system (e.g., key presses for T1 and T2). According to their strategic response deferment model, different lock out strategies are adopted in specific situations to permit performance of T1 and T2 in the manner requested. Whether the response-selection bottleneck is due to a structural limitation on information processing or a strategy adopted to satisfy task demands is an issue that remains to be resolved.
According to response-selection bottleneck accounts of the PRP effect, whether structural or strategic, response selection for T2 does not begin until that for T1 is completed. However, several recent studies have shown cross-talk effects between T1 and T2 that imply that the T2 response is activated before the response for T1 is selected. Hommel (1998) had subjects make a left or right key press to the color of a red or green rectangle for T1 and say “red” or “green” to the letter S or H for T2. RT for both tasks showed correspondence effects at short SOAs, with the response for each task being faster when the color-naming response for T2 corresponded to the color for T1. Lien and Proctor (2000) obtained similar results when T1 involved left-right key presses with the left hand to low or high pitch tones and T2 left-right key presses with the right hand to left-right arrow directions. Also, Logan and Schulkind (2000) reported correspondence effects for the categories of T1 and T2 stimuli for a variety of tasks. For example, when both tasks required letter-digit classifications with left-right key presses on the left and right hands, respectively, RT was shorter when the two stimuli were from the same category (e.g., letters) than when they were not. The fact that, in all studies, the correspondence effects are evident in RT1, as well as RT2, implies that the stimulus for T2 is translated into response activation prior to T1 response selection. Hommel has proposed that such translation of stimulus information into response activation is automatic, with the bottleneck being only in the final decision about which response to make for each task.
Stop Signals
A goal may change during the course of action selection so that the action being selected is no longer relevant. Such situations have been studied in the stop-signal paradigm (Logan, 1994). In this paradigm, a choice-reaction task is administered, but a stop signal occurs at a variable interval after the imperative stimulus on occasional trials to indicate that a response should not be made. Of concern is whether the subject is able to inhibit the response for the choice task. The response is more likely to be inhibited the shorter the interval between the go and stop signals and the longer the choice RT.
Performance on the stop-signal task has been interpreted in terms of a stochastic race model: The go process and stop process engage in a race. The response is executed if the go process finishes before the stop process and is inhibited if the stop process finishes first. This model predicts many features of the results obtained in the stop-signal task, including the probability that the response will be inhibited as a function of go RT and stop-signal delay. The race model has been applied to a variety of stimulus and response modes, suggesting that it captures basic principles of action inhibition. However, it does not provide a detailed account of the processes underlying performance of specific tasks.
Logan and Irwin (2000) compared the processes involved in inhibiting left-right key presses and left-right eye movements. Subjects responded to peripheral left-right stimuli or central left-right pointing brackets, with hand movements or eye movements, using a compatible or incompatible S-R mapping. Estimates of stop-signal RT for hand movements were similar for the two stimulus types and mappings. Stopsignal RT for eye movements was shorter than that for the hands, being shortest for the condition in which a compatible movement was made to a peripheral stimulus. These results suggest that the inhibition processes for hand and eye movements are different, although they follow the same basic principles.
Research has focused on trying to identify the point of no return, or the stage beyond which the response cannot be stopped. De Jong, Coles, Logan, and Gratton (1990) examined this issue using left-right squeezing responses (to a criterion) to measure partial responses, the lateralized readiness potential (LRP) to measure central response activation, and the electromyogram (EMG) to measure muscle activation. LRP, EMG, and squeeze activity were found to occur on stop-signal trials for which the response was successfully inhibited (i.e., did not reach criterion), which they interpreted as suggesting that no stage of response preparation is ballistic. However, Osman, Kornblum, and Meyer (1986) argued that the point of no return should be defined as the point at which the response cannot be stopped from beginning. Using this criterion, the partial squeezes in De Jong et al.’s study are cases of unsuccessful inhibition, indicating that muscle activation is the point of no return. The evidence in De Jong et al.’s study favored two inhibitory mechanisms: Inhibition of central activation processes was implicated because the LRP was truncated on successful stop trials, but several findings suggested that there was also a more peripheral mechanism of inhibition that affected the transmission of activation from central to peripheral structures.
Changes in Action Selection With Practice
Choice RT decreases with practice at a task, with equivalent amounts of practice producing larger changes earlier in practice than later. Teichner and Krebs (1974) reviewed numerous studies of visual choice reactions and concluded that the stage of processing that benefits most from practice is response selection. Newell and Rosenbloom (1981) proposed that the changes in RT with practice follow a power function:
where N is the number of practice trials, B is RT on the first trial, and is the learning rate. The power function has come to be regarded as a law to which any model that is intended to explain practice effects must conform. Although the power law provides a good description of changes in RT with practice averaged across subjects, Heathcote, Brown, and Mewhort (2000) contend that it does not fit the functions for individual performers adequately. They demonstrated that exponential functions provided better fits than power functions to the data for individuals in 40 data sets, and proposed a new exponential law of practice.
Beginning with Merkel (1885), several investigators have shown that the slope of the Hick-Hyman function decreases with practice (e.g., Hyman, 1953; Mowbray & Rhoades, 1959). Seibel (1963) used all combinations of 10 lights assigned directly to 10 keys. After more than 75,000 trials had been performed, the RT for the 1,023-alternative task was only approximately 25 ms slower than that for a 31-alternative task. Practice also is typically more beneficial for incompatible than compatible mappings. However, SRC effects do not disappear even with considerable practice (Dutta & Proctor, 1992; Fitts & Seeger, 1953).
Proctor and Dutta (1993) had subjects perform two-choice tasks for 10 blocks of 42 trials each. In the critical conditions, they performed with the hands uncrossed and crossed in alternate blocks. Whether compatible or incompatible, when the spatial mapping of left-right stimulus locations to leftright response locations remained constant, there was no cost associated with alternating the hand placements: Overall RT and changes with practice with the alternating placements were comparable to those of subjects who practiced with the same hand placement for all blocks. In contrast, when the mapping of stimulus to response locations was switched between blocks so that the same hand was used to respond to a stimulus when the hands were crossed or uncrossed, there was a substantial cost for participants who alternated hand placements compared to those who did not. These results imply that the S-R associations that are strengthened through practice involve spatial response codes.
Practice with an incompatible spatial mapping alters the influence of stimulus location on performance when location becomes irrelevant to the task. Proctor and Lu (1999) had subjects perform a two-choice task for 3 days using an incompatible spatial mapping. On the 4th day, they performed a task for which stimulus location was irrelevant. For this task, the Simon effect was reversed, with RT faster for noncorresponding responses. Tagliabue, Zorzi, Umiltà, and Bassignani (2000) found a similar effect of prior practice with an incompatible mapping and showed it to be present even when subjects were tested a week later. Thus, a limited amount of practice produces new spatial S-R associations that persist at a sufficient strength to override the preexisting associations between corresponding locations.
Nissen and Bullemer (1987) demonstrated that when the trials in a compatibly mapped four-choice spatial reaction task follow a sequence that repeats regularly (every 10 trials in their study), performance improves more with practice than when the trial order is random. Considerable effort has been devoted subsequently to determining whether this sequence learning is implicit or explicit, and to examining the nature of what is learned. Across four experiments, they varied whether the stimuli were spatial or symbolic and whether the responses were spatial or symbolic. Their results showed that the effect of sequence repetition and structure on performance was much stronger for spatial sequences than for symbolic sequences, regardless of whether the stimulus or response set was involved. Koch and Hoffmann also speculated that learning of the response sequence is greater for incompatible S-R mappings (e.g., random mappings of digits to response locations) than for compatible S-R mappings. Regardless, they emphasized, “the selective impact of S-R compatibility on learning stimulus and response sequences in SRT [serial reaction tasks] seems to us an important issue . . . that has not received much attention” (p. 879).
Applications
Contemporary research on action selection has its roots in display-control design issues. Paul Fitts, who formalized the concept of SRC and conducted much of the groundbreaking research on action selection, was the founder of what is now the Fitts Human Engineering Division of the Armstrong Laboratory of the U.S. Air Force and made many contributions to human factors. Although most of the research on action selection has been basic in nature, the results obtained from this research are of considerable relevance to applications involving interface design. It is widely accepted that a userfriendly design must adhere to principles of action selection in general and SRC in particular (see Andre & Wickens, 1990).
In a classic study of stove configurations, Chapanis and Lindenbaum (1959) evaluated four control-burner arrangements (see Figure 11.6a). The experimenter demonstrated the individual pairings of burners to controls for one of the four stoves, and then instructed subjects to push the assigned control to the burner that was lit. Subjects showed shorter RT for Design 1 than Designs 2–4, for which RT did not differ initially. Furthermore, no errors were made for the mappings of Design 1, whereas the overall error rate was 6%, 10%, and 11% for Designs 2–4, respectively. Practice significantly reduced RT and errors for Design 2 compared to Designs 3 and 4, but performance was still worse than with Design 1. When naive subjects were asked which control-burner configuration was the best, most selected Design 1. However, they were equally divided about whether Design 2, 3, or 4 was second best. Thus, although after practice performance was better with Design 2 than Designs 3 and 4, naive subjects did not anticipate this difference.

In a more recent study, Payne (1995) asked naive subjects to rank from easiest to hardest the four mappings of a fourchoice SRC task in which the inner or outer pairs are mapped compatibly or incompatibly (Duncan, 1977; see Figure 11.6b). He compared the subjective rankings to RTmeasures obtained by Duncan. Similar to the results of the stove study, subjects had little difficulty identifying Design 4 as the easiest mapping because it was a direct mapping. However, more subjects rated Design 1 (in which both inner and outer pairs were mapped incompatibly) as being harder than Designs 2 and 3 (in which only one pair was mapped compatibly and the other incompatibly), even though actual performance was second best on Design 1.
The deleterious effect of mixed mappings illustrated in Duncan’s (1977) study, as well as in that of Shaffer (1965), discussed in the “Task Switching” section, indicates that the context in which the display-control configuration is placed affects performance. Most compatibility studies evaluate performance of specific S-R mappings in isolation. However, when more than one S-R mapping is used, the benefit that one might expect from a compatible mapping is not always evident. Moreover, if two tasks must be performed simultaneously, it may be easier to perform them together when they have the same incompatible mapping than when one has a compatible mapping and the other does not (Duncan, 1979). Andre and Wickens (1990) refer to this benefit as one of global consistency and note that it may sometimes be more important than local compatibility.
Because the amount of experience with specific S-R mappings is a major factor in efficiency of action selection, designers must take into account the stereotypic behavior of the specific population for whom a product or system is being designed. For example, from years of experience, Americans are more likely to flick a light switch down when they intend to turn a light off, whereas Englishmen are more likely to flick the switch up. One of the most widely studied population stereotypes is that of direction of motion. Operators of systems that require control of direction of motion must make decisions regarding which direction to move a control in order to move the system or display indicator in a particular direction. Across populations of individuals, many arrangements show preferred relations between the direction of control action and outcome of system output (see Loveless, 1962).
Obviously, when a linear control is in the same orientation as a linear display, the stereotype is to expect the display to move in the same direction as the control. More interesting, when a linear display is oriented perpendicularly to the linear control, right is paired with upward movement and left is paired with downward movement. With control knobs, clockwise rotation tends to be associated with up or right movements (see Hoffman, 1990a, 1990b). In addition, there is a stereotype, called Warrick’s principle, that the display is expected to move in the same direction as the part of the control that is nearest to the display. With a vertical display, a clockwise rotation would be preferred if the control were located to the right of the display and a counter-clockwise rotation if it were located to the left.
Hoffman (1990a, 1990b) evaluated the relative strength of the stereotypes for two- and three-dimensional display and control relationships. He found that different populations (in this case, engineers and psychologists) differed in their preference. Engineers were more likely to follow Warrick’s principle, most likely because it has a mechanical basis, whereas psychologists tended to follow the stereotype of preferring clockwise for up and right movement. This difference emphasizes not only that the specific experience of individuals is important, but also that the preferred relations are based on the individual’s mental model for the task.
Summing Up
Action selection is an important part of behavior inside and outside of the laboratory because choices among alternative actions are required in virtually all situations. Action selection has been a topic of interest in human experimental psychology since Donders’s (1868/1969) seminal work, with contemporary research on the topic being at the forefront of the cognitive revolution in the 1950s. S-R compatibility, which is the quintessential action-selection topic, saw a surge in research in the 1990s, with significant advances made in the development of theoretical frameworks for explaining a variety of phenomena in terms of common mechanisms. As we move into the twenty-first century, the range of tasks and environments in which compatibility effects play a significant role, and the significant insights these effects provide regarding human performance, is only now coming to be fully appreciated.
Bibliography:
- Adam, J. J., Boon, B., Paas, F. G. W. C., & Umiltà, C. (1998). The up-right/down-left advantage for vertically oriented stimuli and horizontally oriented responses: A dual-strategy hypothesis. Journal of Experiment Psychology: Human Perception and Performance, 24, 1582–1597.
- Allport, A., Styles, E. A., & Hsieh, H., (1994). Shifting intentional set: Exploring the dynamic control of tasks. In C. Umiltà & M. Moscovitch (Eds.), Attention and performance: Vol. 15. Conscious and nonconscious information processing (pp. 421–452). Cambridge, MA: MIT Press.
- Andre, A. D., & Wickens, C. D. (1990). Display-control compatibility in the cockpit: Guidelines for display layout analysis. (ARL-90-12/NASA-A3I-90-1). Savoy, IL: University of Illinois, Aviation Research Laboratory.
- Baldo, J. V., Shimamura, A., & Prinzmetal, W. (1998). Mapping symbols to response modalities: Interference effects on Strooplike tasks. Perception & Psychophysics, 60, 427–437.
- Bertelson, P. (1961). Sequential redundancy and speed in a serial two-choice responding task. Quarterly Journal of Experimental Psychology, 13, 90–102.
- Bertelson, P. (1967). The time course of preparation. Quarterly Journal of Experimental Psychology, 19, 272–279.
- Broadbent, D. E. (1958). Perception and communication. Oxford, England: Pergamon Press.
- Campbell, K. C., & Proctor, R. W. (1993). Repetition effects with categorizable stimulus and response sets. Journal of Experimental Psychology: Learning, Memory, and Cognition, 19, 1345–1362.
- Chapanis, A., & Lindenbaum, L. E. (1959). A reaction time study of four control-display linkages. Human Factors, 1(4), 1–7.
- Cho, Y. S., & Proctor, R. W. (2001). Effect of an initiating action on the up-right/down-left advantage for vertically arrayed stimuli and horizontally arrayed responses. Journal of Experiment Psychology: Human Perception and Performance, 27, 472–484.
- Cohen, J. D., Dunbar, K., & McClelland, J. L. (1990). On the control of automatic processes: A parallel distributed processing account of the Stroop effect. Psychological Review, 97, 332–361.
- Cooper, L. A., & Shepard, R. N. (1973). Chronometric studies of the rotation of mental images. In W. G. Chase (Ed.), Visual information processing (pp. 75–176). New York: Academic Press.
- Dalrymple-Alford, E. C., & Budayr, B. (1966). Examination of some aspects if the Stroop color-word test. Perceptual and Motor Skills, 23, 1211–1214.
- De Jagger, J. J. (1970). Reaction time and mental processes (J. Brozek & M. S. Sibinga, Translators). Nieuwkoop, The Netherlands: B. De Graf. (Original work published 1865)
- De Jong, R., Coles, M. G. H., Logan, G. D., & Gratton, G. (1990). Searching for the point of no return: The control of response processes in speeded choice reaction performance. Journal of Experimental Psychology: Human Perception and Performance, 16, 164–182.
- De Jong, R., Liang, C.-C., & Lauber, E. (1994). Conditional and unconditional automaticity: A dual-process model of effects of spatial stimulus-response compatibility. Journal of Experimental Psychology: Human Perception and Performance, 20, 731–750.
- DeSchepper, B., & Treisman, A. (1996). Visual memory for novel shapes: Implicit coding without attention. Journal of ExperimentalPsychology:Learning,Memory,andCognition,22,27–47.
- Donders, F. C. (1969). On the speed of mental processes. In W. G. Koster (Ed.), Acta Psychologica, 30: Vol. 2. Attention and Performance (pp. 412–431). Amsterdam: North-Holland. (Original work published 1868)
- Dosher, B. A. (1979). Empirical approaches to information processing: Speed-accuracy tradeoff functions or reaction time—A reply. Acta Psychologica, 43, 347–359.
- Duncan, J. (1977). Response selection rules in spatial choice reaction tasks. In S. Dornic (Ed.). Attention and performance: Vol. 6 (pp. 49–71). Hillsdale, NJ: Erlbaum.
- Duncan, J. (1979). Divided attention: The whole is more than the sum of the parts. Journal of Experimental Psychology: Human Perception and Performance, 5, 216–228.
- Durgin, F. H. (2000). The reverse Stroop effect. Psychonomic Bulletin & Review, 7, 121–125.
- Dutta, A., & Proctor, R. W. (1992). Persistence of stimulus-response compatibility effects with extended practice. Journal of Experimental Psychology: Learning, Memory, and Cognition, 18, 801–809.
- Eimer, M. (1998). The lateralized readiness potential as an on-line measure of central response activation processes. Behavior Research Methods, Instruments, & Computers, 30, 146–156.
- Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target letter in a nonsearch task. Perception & Psychophysics, 16, 143–149.
- Eriksen, C. W., & Schultz, C. W. (1979). Information processing in visual search: A continuous flow conception and experimental results. Perception & Psychophysics, 25, 249–263.
- Falkenstein, M., Hohnsbein, J., & Hoormann, J. (1994). Effects of choice complexity on different subcomponents of the late positive complex of the event-related potential. Electoencephalography and Clinical Neurophysiology, 92, 148–160.
- Fitts, P. M., & Deininger, R. L. (1954). S-R compatibility: Correspondence among paired elements within stimulus and response codes. Journal of Experimental Psychology, 48, 483–492.
- Fitts, P. M., & Posner, M. I. (1967). Human performance. Belmont, CA: Brooks/Cole.
- Fitts, P. M., & Seeger, C. M. (1953). S-R compatibility: Spatial characteristics of stimulus and response codes. Journal of Experimental Psychology, 46, 199–210.
- Fox, E. (1995). Negative priming from ignored distractors in visual selection: A review. Psychonomic Bulletin and Review, 2, 145–173.
- Gratton, G., Coles, M. G. H., Sirevaag, E. J., Eriksen, C. W., & Donchin, E. (1988). Pre- and poststimulus activation of response channels:Apsychophysiologicalanalysis.JournalofExperimental Psychology: Human Perception and Performance, 14, 331–344.
- Guiard, Y. (1983). The lateral coding of rotations: A study of the Simon effect with wheel-rotation responses. Journal of Motor Behavior, 15, 331–342.
- Heathcote, A., Brown, S., & Mewhort, D. J. K. (2000). The power law repealed: The case for an exponential law of practice. Psychonomic Bulletin & Review, 7, 185–207.
- Hedge, A., & Marsh, N. W. A. (1975). The effect of irrelevant spatial correspondences on two-choice reaction time. Acta Psychologica, 39, 427–439.
- Heister, G., Schroeder-Heister, P., & Ehrenstein, W. H. (1990). Spatial coding and spatio-anatomical mapping: Evidence for a hierarchical model of spatial stimulus-response compatibility. In R. W. Proctor & T. G. Reeve (Eds.), Stimulus-response compatibility: An integrated perspective (pp. 117–143). Amsterdam: North-Holland.
- Helmholtz, H. (1850). Messungen über den zeitlichen Verlauf der Zuckung animalischer Muskeln und die Fortpflanzungsgeschwindigkeit der Reizung in den Nerven [Measurement of the time course of the muscles and the transmission speed of nerve stimulation]. Archiv fürAnatomie und Phsysiologie, 276–364.
- Helmholtz, H. (1867). Mittheilung, betreffend Versuche über die Fortpflanzungsgeschwindigkeit der Reizung in den motorischen Nerven des Menschen, welche Herr. N. Baxt aus Petersburg im Physiologischen Laboratorium zu Heidelberg ausgeführt hat [Communication concerning experiments about the transmission speed of stimulation in the motor nerves of humans, which Mr. N. Baxt from Petersburg in the Physiology Laboratories at Heidelberg has performed]. Monatsberichte der Akademie der Wissenschaften zu Berlin, 29, 228–234.
- Hick, W. E. (1952). On the rate of gain of information. Quarterly Journal of Experimental Psychology, 4, 11–26.
- Hoffman, E. R. (1990a). Strength of component principles determining direction-of-turn stereotypes for horizontally moving displays. Proceedings of the Human Factors Society 34th Annual Meeting, 457–461.
- Hoffman, E. R. (1990b). Strength of component principles for direction-of-turn stereotypes of three-dimensional display/control arrangements. Proceedings of the Human Factors Society 34th Annual Meeting, 462–466.
- Hommel, B. (1993a). Inverting the Simon effect with intention: Determinants of detection and extent of effects of irrelevant spatial information. Psychological Research, 55, 270–279.
- Hommel, B. (1993b). The role of attention for the Simon effect. Psychological Research, 55, 208–222.
- Hommel, B. (1998). Automatic stimulus-response translation in dual-task performance. Journal of Experimental Psychology: Human Perception and Performance, 24, 1368–1384.
- Hommel, B., & Prinz, W. (Eds.). (1997). Theoretical issues in stimulus-response compatibility.Amsterdam: North-Holland.
- Hyman, R. (1953). Stimulus information as a determinant of reaction time. Journal of Experimental Psychology, 45, 188–196.
- Hyman, R., & Umiltà, C. (1969). The information hypothesis and non-repetitions. Acta Psychologica, 30, 37–53.
- Iacoboni, M., Woods, R. P., & Mazziotta, J. C. (1998). Bimodal (auditory and visual) left frontoparietal circuitry for sensorimotor integration and sensoimotor learning. Brain, 121, 2135–2143.
- Jastrow, J. (1890). The time-relations of mental phenomena. New York: N.D.C. Hodges.
- Jensen, A. R. (1980). Chronometric analysis of intelligence. Journal of Social Biological Structure, 3, 103–122.
- Jersild, A. T. (1927). Mental set and shift. Archives of Psychology, 14, (Whole No. 89).
- Kantowitz, B. H., & Sanders, M. S. (1972). Partial advance information and stimulus dimensionality. Journal of Experimental Psychology, 92, 412–418.
- Koch, I., & Hoffmann, J. (2000). The role of stimulus-based and response-based spatial information on sequence learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 863–882.
- Kornblum, S. (1967). Choice reaction time for repetitions and nonrepetitions. Acta Psychologica, 27, 178–187.
- Kornblum, S. (1968). Serial-choice reaction time: Inadequacies of the information hypothesis. Science, 159, 432–434.
- Kornblum, S., Hasbroucq, T., & Osman, A. (1990). Dimensional overlap: Cognitive basis for stimulus-response compatibility—A model and taxonomy. Psychological Review, 97, 253–270.
- Kornblum, S., & Lee, J.-W. (1995). Stimulus-response compatibility with relevant and irrelevant stimulus dimension that do and do not overlap with the response. Journal of Experimental Psychology: Human Perception and Performance, 21, 855–875.
- Kvälseth, T. O. (1980). An alternative to the Hick-Hyman’s and Sternberg’s Law. Perceptual and Motor Skills, 50, 1281–1282.
- Kvälseth, T. O. (1989). Longstreth et al.’s reaction time model: Some comments. Bulletin of the Psychonomic Society, 27, 358–360.
- Lamberts, K., Tavernier, G., & D’Ydewalle, G. (1992). Effects of multiple reference points in spatial stimulus-response compatibility. Acta Psychologica, 79, 115–130.
- Leonard, J. A. (1958). Partial advance information in a choice reaction task. British Journal of Psychology, 49, 89–96.
- Lien, M.-C., & Proctor, R. W. (2000). Multiple spatial correspondence effects on dual-task performance. Journal of Experimental Psychology: Human Perception and Performance, 26, 1260–1280.
- Lippa, Y. (1996). A referential-coding explanation for compatibility effects of perceptually orthogonal stimulus and response dimensions. Quarterly Journal of Experimental Psychology, 49A, 950–971.
- Lippa, Y., & Adam, J. J. (2001). An explanation of orthogonal S-R compatibility effects that vary with hand or response position: The end-state comfort hypothesis. Perception & Psychophysics, 63, 156–174.
- Logan, G. D. (1994). On the ability to inhibit thought and action: A users’ guide to the stop signal paradigm. In D. Dagenbach & T. H. Carr (Eds.), Inhibitory processes in attention, memory, and language (pp. 189–239). San Diego, CA: Academic Press.
- Logan, G. D., & Irwin, D. E. (2000). Don’t look! Don’t touch! Inhibitory control of hand and eye movements. Psychonomic Bulletin & Review, 7, 107–112.
- Logan, G. D., & Schulkind, M. D. (2000). Parallel memory retrieval in dual-task situations: Semantic memory. Journal of Experimental Psychology: Human Perception and Performance, 26, 1072–1090.
- Longsteth, L. E. (1988). Hick’s law: Its limit is 3 bits. Bulletin of the Psychonomic Society, 26, 8–10.
- Longstreth, L. E., El-Zahhar, N., & Alcorn, M. B. (1985). Exceptions to Hick’s Law: Explorations with a response duration measure. Journal of Experimental Psychology: General, 114, 41–74.
- Los, S. A. (1996). On the origin of mixing costs: Exploring information processing in pure and mixed blocks. Acta Psychologica, 94, 145–188.
- Loveless, N. E. (1962). Direction-of-motion stereotypes: A review. Ergonomics, 5, 357–383.
- Lowe, D. G. (1979). Strategies, context and the mechanisms of response inhibition. Memory & Cognition, 7, 382–389.
- Lu,C.-H.,&Proctor,R.W.(1995).Theinfluenceofirrelevantlocation information on performance:Areview of the Simon effect and spatial Stroop effects. Psychonomic Bulletin & Review, 2, 174–207.
- Lu, C.-H., & Proctor, R. W. (2001). Influence of irrelevant information on human performance: Effects of S-R association strength and relative timing. Quarterly Journal of Experimental Psychology, 54A, 95–136.
- MacLeod, C. M. (1991). Half a century of research on the Stroop effect:Anintegrativereview.PsychologicalBulletin,109,163–203.
- Mangun, G. R., Hopfinger, J. B., & Heinze, H.-J. (1998). Integrating electrophysiology and neuroimaging in the study of human cognition. Behavior Research Methods, Instruments, & Computers, 30, 118–130.
- May, C. P., Kane, M. J., & Hasher, L. (1995). Determinants of negative priming. Psychological Bulletin, 118, 35–54.
- McClelland, J. L. (1979). On the time relations of mental processes: A framework for analyzing processes in cascade. Psychological Review, 88, 375–407.
- Merkel, J. (1885). Die zeitliche Verhaltnisse de Willenstatigkeit [The temporal relations of activities of the will]. Philosophische Studien, 2, 73–127.
- Meyer, D. E., & Kieras, D. E. (1997). A computational theory of executive cognitive processes and multiple-task performance: Part 2. Accounts of psychological refractory-period phenomena. Psychological Review, 104, 749–791.
- Michaels, C. F., & Schilder, S. (1991). Stimulus-response compatibilities between vertically oriented stimuli and horizontally oriented responses. Perception & Psychophysics, 49, 342–348.
- Miller, J. (1982). Discrete versus continuous stage models of human information processing: In search of partial output. Journal of Experimental Psychology: Human Perception and Performance, 8, 273–296.
- Miller, J. (1988). Discrete and continuous models of human information processing: Theoretical distinctions and empirical results. Acta Psychologica, 67, 191–257.
- Mordkoff, J. T. (1996). Selective attention and internal constraints: There is more to the flanker effect than biased contingencies. In A. F. Kramer, M. G. H. Coles, & G. D. Logan (Eds.), Converging operationsinthestudyofvisualselectiveattention(pp.483–502). Washington D C:American PsychologicalAssociation.
- Mowbray, G. H., & Rhoades, M. V. (1959). On the reduction of choice reaction times with practice. Quarterly Journal of Psychology, 11, 16–23.
- Neely, J. H. (1977). Semantic priming and retrieval from lexical memory: Roles of inhibitionless spreading activation and limited-capacity attention. Journal of Experimental Psychology: General, 106, 226–254.
- Neill, W. T., & Valdes, L. A. (1992). Persistence of negative priming: Steady state or decay? Journal of Experimental Psychology: Learning, Memory, and Cognition, 18, 565–576.
- Newell, A., & Rosenbloom, P. S. (1981). Mechanisms of skill acquisition and the law of practice. In J. R. Anderson (Ed.), Cognitive skills and their acquisition (pp. 1–55). Hillsdale, NJ: Erlbaum.
- Nicoletti, R., Anzola, G. P., Luppino, G., Rizzolatti, G., & Umiltà, C. (1982). Spatial compatibility effects on the same side of the body midline. Journal of Experimental Psychology: Human Perception and Performance, 8, 644–673.
- Nissen, M. J., & Bullemer, P. (1987). Attentional requirements of learning: Evidence from performance measures. Cognitive Psychology, 19, 1–32.
- Osman, A., Kornblum, S., & Meyer, D. E. (1986). The point of no return in choice reaction time: Controlled and ballistic stages of response preparation. Journal of Experimental Psychology: Human Perception and Performance, 12, 243–258.
- Osman, A., Lou, L., Muller-Gethmann, H., Rinkenauer, G., Mattes, S., & Ulrich, R. (2000). Mechanisms of speed-accuracy tradeoff: Evidence from covert motor processes. Biological Psychology, 51, 173–199.
- Pachella, R. G. (1974). The interpretation of reaction time in information-processing research. In B. H. Kantowitz (Ed.), Human information processing: Tutorials in performance and cognition (pp. 41–82). Hillsdale, NJ: Erlbaum.
- Park, J., & Kanwisher, N. (1994). Negative priming for spatial locations: Identity mismatching, not distractor inhibition. Journal of Experimental Psychology: Human Perception and Performance, 20, 613–623.
- Pashler, H. (1998). The psychology of attention. Cambridge, MA: MIT Press.
- Pashler, H., & Baylis, G. (1991). Procedural learning: 2. Intertrial repetition effects in speeded choice task. Journal of Experimental Psychology: Learning, Memory, and Cognition, 17, 33–48.
- Payne, S. J. (1995). Naïve judgments of stimulus-response compatibility. Human Factors, 37, 495–506.
- Posner, M. I., Klein, R., Summers, J., & Buggie, S. (1973). On the selection of signals. Memory & Cognition, 1, 2–12.
- Prinz, W. (1997). Why Donders has led us astray. In B. Hommel & W. Prinz (Eds.), Theoretical issues in stimulus-response compatibility (pp. 247–267). Amsterdam: North-Holland.
- Proctor, R. W., & Dutta, A. (1993). Do the same stimulus-response relations influence choice reactions initially and after practice? Journal of Experimental Psychology: Learning, Memory, and Cognition, 19, 922–930.
- Proctor, R. W., & Lu, C.-H. (1999). Processing irrelevant location information: Practice and transfer effects in choice-reaction tasks. Memory & Cognition, 27, 63–77.
- Proctor, R. W., & Reeve, T. G. (1985). Compatibility effects in the assignment of symbolic stimuli to discrete finger response. Journal of Experimental Psychology: Human Perception and Performance, 11, 623–639.
- Proctor, R. W., & Reeve, T. G. (1986). Salient-feature coding operationsinspatialprecuingtasks.JournalofExperimentalPsychology: Human Perception & Performance, 12, 277–285.
- Proctor, W., & Reeve, T. G. (Eds.). (1990). Stimulus-response compatibility:Anintegrated perspective.Amsterdam:North-Holland.
- Proctor, W., & Vu, K.-P. L. (2002). Eliminating, magnifying, and reversing spatial compatibility effects with mixed location-relevant and irrelevant trials. In W. Prinz & B. Hommel (Eds.), Common mechanism sinperception andaction: Attention and performance, XIX (pp. 443–473). New York: Oxford University Press.
- Reeve, G. T., & Proctor, R. W. (1984). On the advance preparation of discrete finger responses. Journal of Experimental Psychology: Human Perception and Performance, 10, 541–553.
- Riggio, L., Gawryszewski, L. G., & Umiltà, C. (1986). What is crossed in crossed-hand effects? Acta Psychologica, 62, 89–100.
- Rogers, R. D., & Monsell, S. (1995). Cost of a predictable switch between simple cognitive tasks. Journal of Experimental Psychology: General, 124, 207–231.
- Rosenbloom, P. S., & Newell, A. (1987). An integrated computational model of stimulus-response compatibility and practice. In H. Bower (Ed.), The psychology of learning and motivation (Vol. 21, pp. 1–52). San Diego, CA: Academic Press.
- Roswarski, T. E., & Proctor, R. W. (1996). Multiple spatial codes and temporal overlap in choice-reaction tasks. Psychological Research, 59, 196–211.
- Roswarski, T. E., & Proctor, R. W. (2000). Auditory stimulusresponse compatibility: Is there a contribution of stimulus-hand correspondence? Psychological Research, 63, 148–158.
- Rugg, M. D., & Coles, M. G. H. (Eds.). (1995). Electrophysiology of mind: Event-related brain potentials and cognition. Oxford, England: Oxford University Press.
- Sanders, A. F. (1975). The foreperiod effect revisited. Quarterly Journal of Experimental Psychology, 27, 591–598.
- Sanders, A. F. (1998). Elements of human performance. Mahwah, NJ: Erlbaum.
- Sanders, A. F., & Wertheim, A. H. (1973). The relation between physical stimulus properties and the effect of foreperiod duration on reaction time. Quarterly Journal of Experimental Psychology, 25, 201–206.
- Schweickert, R. (1983). Latent network theory: Scheduling of processes in sentence verification and the Stroop effect. Journal of Experimental Psychology: Learning, Memory, and Cognition, 9, 353–383.
- Seibel, (1963). Discrimination reaction time for a 1,023-alternative task. Journal of Experimental Psychology, 66, 215–226.
- Shaffer, L. H. (1965). Choice reaction with variable S-R mapping. Journal of Experimental Psychology, 70, 284–288.
- Smith, E. E. (1968). Choice reaction time: An analysis of the major theoretical positions. Psychological Bulletin, 69, 77–110.
- Soetens, E. (1998). Localizing sequential effects in serial choice reaction time with the information reduction procedure. Journal of Experimental Psychology: Human Perception and Performance, 24, 547–568.
- Sternberg, S. (1969). The discovery of processing stages: Extensions of Donders’ method. In W. G. Koster (Ed.), Attention and performance II. Acta Psychologica, 30, 276–315.
- Sternberg, S. (1998). Discovering mental processing stages: The method of additive factors. In D. Scarborough & S. Sternberg (Eds.), An invitation to cognitive science: Methods, models, and conceptual issues (2nd ed., Vol. 4, pp. 703–863). Cambridge, MA: MIT Press.
- Stoffer, T. H., & Umiltà, C. (1997). Spatial stimulus coding and the focus of attention in S-R compatibility and the Simon effect. In B. Hommel & W. Prinz (Eds.), Theoretical issues in stimulus-response compatibility (pp. 181–208). Amsterdam: North-Holland.
- Stroop, J. R. (1992). Studies of interference in serial verbal reactions. Journal of Experimental Psychology: General, 121, 15–23. (Original work published 1935)
- Tagliabue, M., Zorzi, M., Umiltà, C., & Bassignani, F. (2000). The role of LTM links and STM links in the Simon effect. Journal of Experimental Psychology: Human Perception and Performance, 26, 648–670.
- Teichner, W., & Krebs, M. J. (1974). Laws of visual choice reaction time. Psychological Review, 81, 75–98.
- Telford, C. W. (1931). The refractory phase of voluntary and associative responses. Journal of Experimental Psychology, 14, 1–36.
- Umiltà, C., & Nicoletti, R. (1990). Spatial stimulus-response compatibility. In R. W. Proctor & T. G. Reeve (Eds.), Stimulusresponse compatibility: An integrated perspective (pp. 89–116). Amsterdam: North-Holland.
- Van Zandt, T., Colonius, H., & Proctor, R. W. (2000). A comparison of two response time models applied to perceptual matching. Psychonomic Bulletin & Review, 7, 208–256.
- Vickrey, C., & Neuringer, A. (2000). Pigeon reaction time, Hick’s law, and intelligence. Psychonomic Bulletin & Review, 7, 284–291.
- Vu, K.-P. L., & Proctor, R. W. (2001). Determinants of right-left and top-bottom prevalence for two-dimensional spatial compatibility. Journal of Experimental Psychology: Human Perception and Performance, 27, 813–828.
- Vu, K.-P. L., & Proctor, R. W. (2002). Mixing compatible and incompatible mappings: Elimination, reduction, and enhancement of spatial compatibility effects. Manuscript submitted for publication.
- Wang, H., & Proctor, R. W. (1996). Stimulus-response compatibility as a function of stimulus code and response modality. Journal of Experimental Psychology: Human Perception and Performance, 22, 1201–1217.
- Weeks, D. J., & Proctor, R. W. (1990). Salient-features coding in the translation between orthogonal stimulus and response dimensions. Journal of Experimental Psychology: General, 119, 355–366.
- Weeks, D. J., Proctor, R. W., & Beyak, B. (1995). Stimulus-response compatibility for vertically oriented stimuli and horizontally oriented responses: Evidence for spatial coding. Quarterly Journal of Experimental Psychology, 48, 367–383.
- Welford, A. T. ( 1987). Comment on “Exceptions to Hick’s law: Explorations with a response duration measure” (Longstreth, El-Zahhar, & Alcorn, 1985). Journal of Experimental Psychology: General, 116, 312–314.
- Wickelgren, W. A. (1977). Speed-accuracy tradeoff and information processing dynamics. Acta Psychologica, 41, 67–85.
- Woodworth, R. S. (1938). Experimental psychology. New York: Holt.
- Wundt, W. (1883). Über psychologische Methoden [Over psychological methods]. Philosophische Studien, 10, 1–38.
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality