
Sample Genotype And Phenotype Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
The fundamental distinction between genotype and phenotype was first introduced into biology by Wilhelm Johannsen in 1911 to make explicit a distinction that was implicit in Mendel’s explanation of heredity, and which marked it off from previous theories of inheritance. Mendel found that when he crossed two pure-breeding varieties of garden pea, the first generation of hybrid offspring were indistinguishable from one of the two parental strains, but if these hybrids were crossed with each other, both of the original types appeared in the offspring of these hybrids. So, for example, a pure bred red-flowered crossed with a pure bred white-flowered form produced all red-flowered plants, but these, when interbred with each other produced both red and white-flowered plants.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
In order to explain this extraordinary result, Mendel postulated the existence of internal, unseen ‘factors’ contributed by each parent to the offspring, factors that were causal determinants of the outward appearance of the plants, but whose nature was unaffected by their passage from one generation to the next. So, pure bred white-flowered plants had two white ‘factors’ one contributed by the pollen and one by the egg of their male and female parents, while pure bred red-flowered plants had received two red ‘factors’ from their two parents. The hybrids of red-flowered and white flowered forms would then contain one red and one white ‘factor,’ yet these plants were indistinguishable in appearance from a parent that had two red ‘factors.’ There was then a fundamental distinction between the state of the internal causal ‘factors’ and the outward state of the plant that carried them. Moreover, to explain the reappearance in the next generation of some white-flowered plants, Mendel had to suppose that the temporary coexistence of red and white ‘factors’ in the hybrids had no effect on their nature, but that they could be passed on uncontaminated to the offspring of the hybrids. An offspring of the hybrids that happened to contain two white ‘factors’ was then white like its white grandparent.
This scheme of explanation makes a radical separation between the internal factors that behave according to one set of physical relations in their passage between generations, the laws of heredity, and the external appearance of the organism that results from a different set of physiological relations which determine the consequences to the organism of carrying those factors, the laws of Development. This distinction between the phenomenon of heredity and the phenomenon of development which we usually associate with August Weissman’s separation of the germplasm from the somatoplasm had already been made by Mendel twenty years earlier. Earlier theories of heredity had not made such a separation of inner factors from outward manifestation and had been posed as the problem of the inheritance of the organism’s characters themselves. It was Mendel’s realization that characters are not inherited, they develop. In Johanssen’s terminology the description of an organism’s internal factors that were passed on to it in the gametes from its parents is a specification of the organism’s genotype, while the description of the total physical and behavioral state of the organism is a specification of its phenotype.
1. Description Of Genotypes And Phenotypes
As a result of the discoveries in molecular and cellular biology since Johanssen’s time genotypic and phenotypic descriptions have become more concrete, more detailed and greatly amplified in their scope. Originally genotypes had no independent physical descriptors but were distinguished from each other entirely on the basis of the external phenotypic manifestations and the patterns of similarity in phenotypes of related individuals. ‘Genes’ were simply abstract placeholders of unknown physical composition, inferred entities that could be characterized only by their phenotypic effects.
The modern identification of DNA as the physical material that behaves in heredity in the manner inferred for Mendel’s mysterious ‘factors’ has allowed a precise physical description of the genotype of an organism. It is the complete spatially-ordered sequence of the four alternative nucleotide bases, adenine, cytosine, guanine and thymine contained in the DNA of the organism’s cells. In its most complete form this genotypic description includes not only the DNA in the nucleus of the cell but in certain cellular organelles such as mitochondria or chloroplasts that carry the DNA that is related to their physiological function and that is passed across generational lines. Phenotypic description has also been greatly amplified beyond the external characteristics of individuals. It now includes the gross structure of the various cells of the organism, the physico-chemical description of the various molecules within cells, including their shape and intra-cellular localization, and the network of biochemical reactions in which these molecules participate.
The concepts of phenotype and genotype require a distinction between types and tokens. Phenotype and genotype are, as the words indicate, types, sets of which individual organisms are members, sets delineated by the complete descriptors of those organisms. Any individual member of these type sets is a token of the type. In addition it is necessary to distinguish the descriptors of organisms from the physical object or behavior that is being described. The concepts phenotype and genotype refer to the descriptors. The physical objects that are being described are the phenome and genome of an organism, the actual physical material or behavior of some particular individual and its actual DNA molecules. If we take the definitions literally, then no two organisms belong to the same phenotype because there is always some physical difference in morphology, physiology and behavior between any two organisms, even between identical twins, cloned individuals, or cells that result from asexual reproduction. Moreover, except for the results of twinning, cloning or vegetative reproduction, no two individuals belong to the same genotype, as the process of mutation guarantees some variation in their DNA between any two individuals. The rate of mutation per nucleotide per cell division is of the order of one in a billion, so for mammals, with over a billion nucleotides of DNA making up their genes, even identical twins are likely to have some small difference in total genotype. There is, in addition, an ambiguity in the assignment of an individual to a genotype because mutations occur during the millions of cell divisions that occur in development, so that all cells in the body do not have identical DNA.
In practice phenotypic and genotypic descriptions are not total but partial, referring to some aspect of the genome and phenome that are of particular interest, while ignoring the rest. The partial phenotypic and genotypic descriptions are partial in two senses. First, only some aspect of the morphology or function of the organism is characterized; say the rate of enzymatic splitting of alcohol, a phenotype, and the DNA sequences that code for the enzyme proteins involved in the reaction, a genotype. Second, a decision must be made about the collection of actual partial phenotypes and genotypes that will be regarded as indistinguishable. At the phenotypic level, given the actual variation in enzymatic rates from individual to individual and from time to time, what range of rates will be regarded as belonging to the same phenotypic class? For any phenotypic variable that is continuous, some boundaries of distinguishable phenotypic classes must be laid down in order to make a mapping between phenotype and genotype because genotypes, by their nature, are discrete classes.
The problem of aggregation exists at the genotypic level as well. The relation of DNA differences to differences in the amino acid sequence of a protein is not one-to-one. Because the DNA triplet code is redundant, substitutions of nucleotides in the third position of the triplet often do not change the amino acid that is specified, so for some purposes many different alternative DNA sequences of a gene count as the same genotype. On the other hand, nucleotide substitutions that do not affect the amino acid sequence of a protein may affect the rate of synthesis of that protein and so may, for some questions, be counted as belonging to different genotypes.
The decision about which aspects of total phenotype and genotype are relevant in creating genotypic and phenotypic classes is a deep question of biology that is largely ignored because everything cannot be considered at once. Sometimes biologists act as if the rest of the organism really were constant, sometimes they recognize the variation but claim that it is causally irrelevant to the phenomenon of interest and sometimes they admit its causal relevance but claim that the variation can be treated as background experimental noise that can be averaged out by repeated observations on a large class of individuals. These decisions are usually made either for the sake of observational convenience without a coherent rationale or from a commitment to a reductionist view of the organism that asserts that an understanding of the whole system can be obtained by breaking it down into ‘obvious’ small components to be studied in isolation.
2. Relationship Between Genotype And Phenotype
It is clearly true that organisms do indeed consist of a collection of quasi-independent genotypic and phenotypic subsystems, within which there are strong interactions and between which there are very weak interactions. But the boundaries of these genetic and phenotypic subsystems can only be found by actual experiment and differ from function to function. The hand and eye are, as far as is known, developmentally independent systems for which different gene combinations are relevant during embryogenesis, but handeye coordination is essential for reaching out and picking up observed objects and so cannot be regarded as phenotypically or genotypically independent in considering this elementary behavior.
The problem for genetics and development is how the genotypic and phenotypic spaces map into each other. It is now certain that Weissman was correct in claiming that the genotype-phenotype mapping is causally asymmetrical. Genotypic differences may cause phenotypic differences during the process of development, but there is no material pathway by which changes in the phenome can result in related changes in the genome. There is no biological inheritance of acquired characters. This causal asymmetry sets the program for developmental, physiological and behavioral genetics which is to discover the mechanisms by which the reading of DNA by the cell machinery is causally efficacious in influencing the phenotypic state of the organism. This is the problem of making the forward mapping of genotype on to phenotype.
There remains, then, the problem of the backward mapping, how the causal knowledge of epigenetic processes can be used to infer the genotype, given the phenotype. Even if a knowledge of the genotype gave perfect information about the phenotype, the inverse would not be true, as discovered by Mendel. The phenomenon of dominance alone makes it impossible to distinguish between heterozygotes and homozygotes from phenotypic information at an arbitrary level. In addition, the same phenotype may appear as a result of changes at different locations in the DNA. There are, for example, more than 200 different nucleotide substitutions known in the genes coding for blood-clotting proteins, any one of which leads to hemophilia. The major difficulty of mapping between genotypes and phenotypes is, however, in the forward direction. The complete DNA sequence of an organism does not contain the information necessary to specify that organism.
3. Genotype, Phenotype, And Environment
First, the outcome of development and the ongoing functioning of an individual is the consequence of a unique interaction between the molecules contained in the fertilized egg, including the DNA, and the sequence of environments in which development occurs. Figure 1 shows the result of a cloning experiment in a plant, Achillea, grown at three different elevations in the Sierra Nevada. Immature plants were sampled from nature, each plant was cut into three pieces, and one piece was replanted at each of the three elevations. The figure shows the height to which the pieces grew in the different environments, each line connecting the three genotypically identical pieces in the three environments. A line, giving the phenotype of a given genotype as a function of different environments is a norm of reaction. The norms of reaction in this case show that there is no genotype with unconditionally better growth and, in fact, there is no way to predict the order of growth of the different genotypes in one environment from their order of growth in another environment. Thus, the phenotype is not only contingent on the environment but in an unpredictable and irregular way.
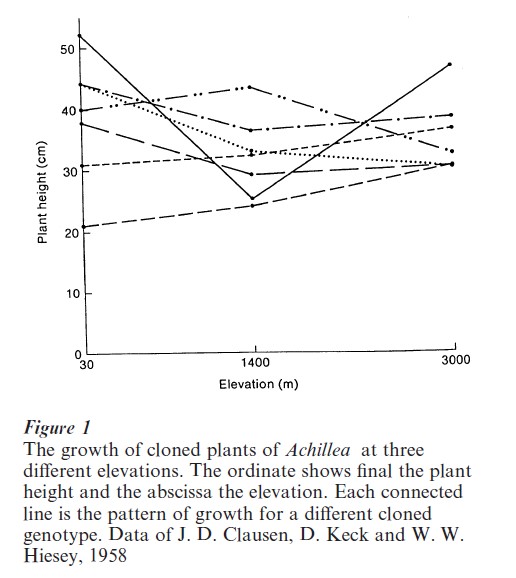
Such irregular and particularistic relations between genotype and phenotype in different environments is characteristic of norms of reaction whenever they have been measured for morphological and behavioral characteristics in experimentally accessible organisms. There are, of course, extreme exceptions; genotypes that result in the same phenotype in any environment and environments that are in no way contingent on genotypes for their effect. Eye color variation among humans is entirely a consequence of genotypic differences, uninfluenced by environment, while regional and social class differences in the phonemic structure of speech have no influence of genetic differences. Moreover, major differences between species are a consequence of genetic differences that are manifest in all environments. There is no environment in which a gorilla could learn to write this research paper. Within species, however, the vast majority of phenotypic variations in morphology, physiology, and behavior are a consequence of particular contingent relations between genotype and phenotype.
4. Developmental Noise
Second, even a complete specification of the genotype and the environment is insufficient to specify the organism. All cells contain a very small number of copies of biologically active molecules, spatially segregated within the cell, so that the number of various reactions per unit time vary greatly from cell to cell and time to time. Moreover, when cells divide they do not distribute the molecules exactly evenly, except for DNA which has a special machinery for guaranteeing its equal distribution. The consequence of these small molecular numbers is that there is random variation from cell to cell in time to the next cell division, in cell size, in cell shape and in the movements of cells during development. These random events, random at the level of thermal and quantum chemical molecular states, can have considerable effect on the formation and behavior of the whole organism. (Goss and Pecoud 1998)
So-called ‘bilateral symmetries’ are not, in fact, symmetrical but vary between sides in an irregular way from individual to individual. The fingerprints on the left hand are not identical, although similar to, those on the right. Such random Developmental noise is a nontrivial source of variation in phenotype. During the development of the central nervous system of a multicellular organism, superimposed on the major regular patterns of neuronal connection are random variations in the growth and connections of specific neuronal pathways, even in organisms like the roundworm Caenorhabditis with a very simple central nervous system. It has yet to be determined how much of the behavioral variation among individuals is a consequence of such random variation in the development of the central nervous system, variation that is inborn, but not genetic.
The environmental and chance contingency of the mapping between genotype and phenotype arises from the nature of organisms as physical objects. They are intermediate in size between particles and planets and they are internally functionally heterogeneous. As a result, unlike simple physical systems like the movement of planets around the sun, their functions are not the consequence of a few strong causal forces whose parameters govern the outcome. Organisms are the nexus of a very large number of individually weakly determining forces subject to noise at all levels. The consequence for the understanding of the physical properties of organisms, including their individual and social behavior, is that we cannot expect any simple and universal generalizations like Newton’s Laws. That is certainly true for the relation between genotype and phenotype.
Bibliography:
- Clausen J D, Keck D, Hiesey W W 1958 Experimental Studies on the Nature of Species Vol. 3: Environmental Responses of Climatic Races of Achillea. Carnegie Institution of Washington Publication No. 581
- Goss P, Pecoud J 1998 Quantitative modeling of stochastic systems in molecular biology using stochastic petri nets. Proceedings of the National Academy of Science USA 95: 6750–5
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality