
Sample Coevolution Of Genes And Culture Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
Cultural transmission can be roughly defined as the transfer of information between individuals by social learning. Genes used to be—and for most species still are—the only means available for the accurate transfer of information across generations. In species where cultural transmission has developed, notably the human, interactions can occur between the two inheritance systems. Gene-culture coevolution refers to the evolutionary phenomena that arise from these interactions.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Quantifying Cultural Evolution
The principles of genetic evolution are well established. By contrast, cultural evolution is more contentious. The information that forms the basis of culture is transmitted by social learning—instruction by, or imitation of, other individuals. This information must somehow reside in the brain, but it is unclear how this is achieved. Nevertheless, just as significant insights into genetic evolution were obtained before DNA was identified as the genetic material, cultural evolution can be studied without waiting for definitive progress in cognitive science.
Central to the modern treatment of cultural evolution is the concept of a ‘meme’ (Dawkins 1976; also ‘culturgen’ of Lumsden and Wilson 1981). The meme is defined, in analogy with the gene, as a unit of information that is transmitted by social learning. Surnames are a good example of a meme that is highly congruent to a gene. Surnames are well-defined units that in many societies are transmitted from father to son, just like a gene on the Y chromosome. However, most memes cannot be delimited as precisely, so that copies of the same meme in two individuals may differ in content. Consider the ambiguity of ‘how to use a personal computer.’ In addition, although the meme is defined as information, it is usual to regard as a meme the more readily observable behavior that the information regularly induces.
Memes also differ from genes in that there is a multiplicity of transmission pathways. ‘Vertical’ transmission from parents to their children and ‘horizontal’ transmission between individuals of the same generation have received the most attention.
The classical, although by no means the first, quantitative study of horizontal transmission is due two rural sociologists, Ryan and Gross (1943), who investigated the spread of hybrid seed corn. This striking technical advance in agriculture was introduced to Iowa farming communities in the late 1920s. Ryan and Gross learned through retrospective interviews when each farmer had adopted this innovation. By the time these interviews were conducted in 1941, adoption was almost complete. The cumulative number of adopters follows an S-shaped curve, which resembles the logistic curve. It is not known whether this resemblance is more than superficial.
The study of vertical transmission begins with Feldman and Cavalli-Sforza (1976). Assume two alternative memes, such as whether or not salty foods are eaten habitually. In general, denote the alternative memes by H and h. Then, the four possible combinations for husband and wife are H×H, H×h, h×H, or h×h. If children acquire their memes by vertical transmission—by social learning from their parents—the four types of families should produce different frequencies of H and h children. More specifically, if the probabilities of an H child in the four types of families are b3, b2, b1, and b0, respectively, then the inequalities b3 ≥b2, b3≥ b1, b2≥ b0 , and b1≥ b0 should hold. A survey of Stanford University students and their parents showed that, for the example of habitual salt use, b3= 60 percent, b2=36 percent, b1=65 percent, and b0= 26 percent, suggesting a fairly strong maternal influence (Cavalli-Sforza and Feldman 1981).
Cultural evolution is said to occur when the frequency (proportion) of a meme in a population changes. Memes are in principle subject to the same evolutionary forces as genes—the analog of mutation, random drift, natural and sexual selection. For example, the meme for habitual salt use may raise mortality and as a result be selected against. On the other hand, an important force acting on memes to change their frequencies, but one not normally applicable to genes, is transmission bias. The spread of hybrid seed corn in the Iowa communities is an example of cultural evolution by transmission bias, since nonadopters are more likely to be converted to adopters than vice versa.
Having briefly described cultural evolution, let us now consider in detail two well-documented instances of an interaction between genetic and cultural evolution. Many more case studies of gene-culture coevolution are discussed in books by Lumsden and Wilson (1981), Boyd and Richerson (1985), and Durham (1991).
2. Adult Lactose Absorption And Drinking The Milk Of Domesticated Animals
Perhaps the paradigmatic example of gene-culture coevolution is the process whereby the genetic ability of adult humans to digest lactose (adult lactose absorption) became associated with their consumption of animal milk.
The fresh milk of domesticated animals, such as goats, camels, and cows, has been an important food in northern Europe and in pastoral societies for about 5,000 to 6,000 years. By contrast, animal milk is not traditionally used by hunter–gatherer and nondairying agricultural groups.
Milk contains a sugar, lactose, which must be digested by the enzyme, lactase, before it can be absorbed in the small intestine. All normal infants have high lactase activity, but during development the majority of individuals experience a drop in lactase activity, which is fully manifest by school age. Hence, adults are of two types—lactose absorbers who have high lactase activity, and lactose malabsorbers who have low lactase activity. Data from families and individuals of mixed ancestry indicate that adult lactose malabsorption is inherited as a simple Mendelian recessive.
When milk is consumed in quantity by a lactose malabsorber, the lactose passes undigested into the colon. Symptoms of intolerance such as severe diarrhea can result. Malabsorbers may also be less efficient at absorbing the calcium in milk.
Many researchers have noted the following pattern. First, populations with high frequencies of absorbers were traditionally dependent on animal milk. Second, populations not known to have used milk have low frequencies of absorbers. The highest frequencies are observed in northern Europe among the Finns, Danes, and Germans. Comparable frequencies are seen among the nomadic tribes of the north African and Arabian deserts, such as the Beja, Tuareg, and Jordanian Bedouin. On the low end are groups like the Japanese, the Pima Indians of Arizona, and the Yoruba of central Africa, none of which have a history of dairying. Although the covariation is impressive, it is not the case that all dairying populations have a high frequency of absorbers, and numerous ‘exceptions’ can be found around the Mediterranean. Hence, populations with a high frequency of absorbers apparently form a subset of those with a history of milk use.
Lactase activity drops at weaning in nonhuman mammals, suggesting that our ancestors were also malabsorbers. This inference and the above observations led to the proposal of the ‘culture-historical’ hypothesis, which posits that lactose absorbers gained a selective advantage when children and adults began to drink the milk of domesticated animals (Simoons 1970, McCracken 1971, Kretchmer 1972). Hence, the frequency of absorbers increased by natural selection in populations that had access to animal milk—or at least in some of them.
A variant of the culture–historical hypothesis is the ‘calcium absorption’ hypothesis (Flatz and Rotthauwe 1973). Whereas the former attributes a general nutritional advantage to the use of milk, the latter claims that calcium was all important. Recall that high frequencies of absorbers are observed in northern Europe. Here, solar radiation is weak, and the calcium in milk may have provided protection from rickets.
A different hypothesis called the ‘reverse cause’ argument suggests that dairying was adopted by precisely those populations that could tolerate lactose. The genetic differentiation of human groups is assumed to have occurred before the invention of dairying by some process unrelated to milk use. In particular, drinking milk is not assumed to have conferred a selective advantage.
The two major proposals, the culture–historical hypothesis and the reverse cause argument, thus differ in the temporal priority given to cultural or to genetic change. (A third possibility is that they occur together, in which case the process would be truly coevolutionary.) However, the prevailing view in modern anthropology is that the genetic differences (if any) among human populations are not the cause of the cultural differences among those populations. In particular, the claim that a preexisting genetic contrast, such as in the prevalence of lactose absorbers, would have produced divergent cultural evolution is anathema. Hence, the reverse cause argument, in straying from dogma, has already acquired one strike against it.
More substantively, Holden and Mace (1997) have recently used phylogenetic comparative methods to show that the cultural evolution of milking probably preceded the genetic evolution of lactose absorption. In a sense, this outcome is anticipated by the fact noted above that a high frequency of absorbers is observed in only some of the dairying populations and in none of the nondairying ones. In addition, this analysis reveals that statistical evidence for the calcium absorption hypothesis is weak. Instead, the parsimonious explanation is that northern European populations are similar in having high frequencies of absorbers because they share a relatively recent common ancestral population. On the other hand, the culture–historical hypothesis is supported by a global phylogenetic analysis.
In the previous section, it was explained how a useful innovation may spread by horizontal transmission. Presumably, drinking the milk of domesticated animals can be regarded as a useful innovation, at least for absorbers. Hence, in societies where domesticated animals were available, as short a time as one generation may have sufficed for dairying to become established. If so, the claim that milk use spread first and lactose absorption evolved later may be valid.
Against this, however, the possibility must be entertained that drinking milk was an unattractive option to a society dominated by lactose malabsorbers. In other words, there may be a genetically determined transmission bias against milk use among malabsorbers. Studies conducted in modern industrial societies reveal little evidence of a clearcut difference in preference for milk between absorbers and malabsorbers. This may be because the availability of many foods, and especially the presence of lactose as an additive, makes it difficult for malabsorbers to associate the symptoms of intolerance with the ingestion of milk. By contrast, in traditional societies where food choices are limited, a difference in preference may be easier to demonstrate.
3. Recessive Hereditary Deafness And Sign Language
The incidence of profound childhood deafness is about one in 1,000. About one half of these cases is hereditary, with recessive genes in particular being responsible for about one third of the total incidence. Estimates of the number of recessive genes range from five to more than 30 depending on the population. Homozygosity for (having two copies of ) any one of these genes results in deafness. Among the nongenetic causes are maternal rubella, neonatal jaundice due to rhesus incompatibility, and otitis media.
Sign languages are the naturally occurring forms of linguistic communication among the profoundly deaf. A sign language is not a collection of gestures. Rather, grammatical rules govern the production of words and the expression of thoughts. There are many sign languages just as there are many spoken languages. Sign languages are natural languages, whose origins cannot be determined and which have existed in the deaf communities possibly for many generations.
When deafness is recessive, an affected child may be born to normal heterozygous (having one copy of a gene for deafness) parents. In fact, about 90 percent of deaf children have hearing parents. A hearing child of hearing parents will learn the spoken language used by the parents. Vertical transmission plays an important role in the transmission of a spoken language across generations. By contrast, a deaf child of hearing parents usually cannot learn the local sign language from the parents, simply because the latter do not know it. Hence, the transmission of a sign language across generations is complicated by the fact that a significant fraction of profound childhood deafness is caused by recessive genes. Aoki and Feldman (1991), therefore, suggest that a sign language is a good example of a meme whose transmission and social distribution are subject to strong, if indirect, genetic constraints.
Another factor that interferes with the vertical transmission of a sign language is natural selection against the deaf. Selection is apparently due, not to higher mortality, but to lower marriage and fertility rates among the deaf. In the 1950s a deaf person in Northern Ireland/or Japan could expect to have about 30 percent as many children as a hearing person. These observations suggest that a sign language cannot persist under vertical transmission alone. This prediction is supported by a simple model, which is presented below to illustrate some of the principles of gene-culture coevolution.
Assume the following: (a) One gene causing deafness segregates in the population. There are two alleles (alternative states of a gene), the normal allele, and the mutant allele that causes deafness when homozygous; (b) Nongenetic causes of deafness are ignored; (c) The deaf suffer a higher mortality than the hearing, which is contrary to fact, but has approximately the same evolutionary effect as lower nuptiality or fertility. The relative viability of a deaf person is ; (d) The recessive allele causing deafness arises by mutation in each generation; (e) Deaf persons tend to marry other deaf persons. The rate of assortative mating is m; (f ) Only the deaf use sign language. Sign language is transmitted vertically from signing parent(s) to deaf children. A deaf child acquires sign language with probability c, kc, or 0 depending on whether there are two, one, or no signing parents. The correspondence with the parameters of Sect. 1 is b3=c, b2=b1=kc, b0=0. When the child is hearing, b3=b2=b1=b0=0.
Assumptions (a)—(e) and assumption (f ) determine the dynamics of the gene for deafness and the meme for sign language, respectively. Together, they constitute a model for the coevolution of genes and culture. When the strong force of natural selection and the weak force of mutation balance each other, the allele causing deafness will exist at a low equilibrium frequency. The dynamics of the frequency of signers, x, can then be approximately described by the following recursion equation, where the prime indicates the value in the next generation.
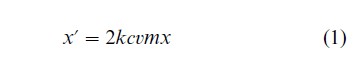
Equation (1) is valid when the mutation rate and the frequency of signers are low, which are realistic assumptions.
Clearly, the frequency of signers will decrease and sign language will be lost if 2kcvm<1. On the other hand, sign language can persist if 2kcvm>1. Estimates of the strength of natural selection and of the rate of assortative mating are available for Japan and Northern Ireland. For example, in Japan in the 1950s, m=62 percent and v=30 percent. Substitution gives 2kcvm =0.37×kc. This is smaller than one, because the vertical transmission parameter, kc, cannot exceed one.
Thus, this simple coevolutionary model suggests that a sign language cannot be maintained in a deaf community by vertical transmission alone. Some linguists invoke horizontal transmission at schools for the deaf, where naive deaf children can learn from classmates who were fortunate enough to have deaf signing parents.
Bibliography:
- Aoki K, Feldman M W 1991 Recessive hereditary deafness, assortative mating, and persistence of a sign language. Theoretical Population Biology 39: 358–72
- Boyd R, Richerson P J 1985 Culture and the Evolutionary Process. University of Chicago Press, Chicago
- Cavalli-Sforza I L, Feldman M W 1981 Cultural Transmission and Evolution. Princeton University Press, Princeton, NJ
- Dawkins R 1976 The Selfish Gene. Oxford University Press, New York
- Durham W H 1991 Coevolution. Stanford University Press, Stanford, CA
- Feldman M W, Cavalli-Sforza L L 1976 Cultural and biological evolutionary processes, selection for a trait under complex transmission. Theoretical Population Biology 9: 238–59
- Flatz G, Rotthauwe H W 1973 Lactose nutrition and natural selection. Lancet 2: 76–7
- Holden C, Mace R 1997 Phylogenetic analysis of the evolution of lactose digestion in adults. Human Biology 69: 605–28
- Kretchmer N 1972 Lactose and lactase. Scientific American 277(April): 71–8
- Lumsden C J, Wilson E O 1981 Genes, Mind, and Culture. Harvard University Press, Cambridge, MA
- McCracken R D 1971 Lactase deficiency: An example of dietary evolution. Current Anthropology 12: 479–517
- Ryan B, Gross N C 1943 The diffusion of hybrid seed corn in two Iowa communities. Rural Sociology 8: 15–24
- Simoons F J 1970 Primary adult lactose intolerance and the milking habit: A problem in biologic and cultural interrelations. II. A culture historical hypothesis. American Journal of Digestive Disease 15: 695–710
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality