
Sample Genetics And Development Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
This research paper describes the major historical misconceptions of the role of genetics in the development of the individual, whether human or nonhuman animal, and also presents our current understanding of the necessarily co-operative roles of genes and environments in the developmental process.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Nature vs. Nurture
Many people believe that behavioral characteristics such as personality or intelligence are determined primarily, if not exclusively, either by heredity or environment. This belief, known as the nature-nurture controversy, has been with us for quite a long period of time and is the cause for much that is sorrowful and tragic in human history (e.g., the notion that some groups are behaviorally inferior or superior to other groups because of hereditary genetic predispositions). The idea that genes are primarily responsible for some traits and that the environment is primarily responsible for other traits is now known to be scientifically invalid. This has been a hard-won scientific insight that has not yet percolated down to the mass of humanity. It is now known that both genes and environments are involved in all traits and that it is not possible to specify their respective weighting or quantitative influence on any trait. These crucial points are documented in the sections that follow.
2. Behavioral Genetics
In a remarkable experiment first conducted by E. C. Tolman (1924) and continued by R. C. Tryon (1942), it was found that it was possible to selectively breed laboratory rats for their ability to learn their way through a maze. Tryon selectively inbred the superior learners among each other and the poor learners among each other. After only seven generations of breeding the brightest rats within each of the brightest litters, and the dullest within each of the dullest litters, the performance of the two groups in learning the specific maze was virtually nonoverlapping, leading Tryon to label the superior line ‘maze-bright’ and the inferior line ‘maze-dull.’ It was as if Tryon had been able, by selective breeding, to create two lines of animals which differed greatly in their general learning ability. That turned out not to be the case, however. When Searle (1949) later compared the ‘bright’ and ‘dull’ lines on a variety of learning and performance tasks, the dull rats were sometimes superior and often equal to the rats from the bright line. Being bright or dull was not a property of the animals, but, rather, was restricted to the particular learning task that served as the basis for selectively breeding the animals. As it turned out, the specifics of the outcome of the breeding program were even further constrained by the manner in which the rats were reared, as will be described in Sect. 3.
Since the time of Tolman’s and Tryon’s breeding experiments, the field of behavioral genetics has been able to show that it is possible to selectively breed animals for differences on any trait, solidifying the idea mentioned earlier that genes are involved in all traits. As will be documented below, the results of behavioral genetics breeding experiments are also dependent on the rearing environment being kept constant from generation to generation (recall that the rearing environment is involved in all traits, too). As can be seen in Table 1, both favorable and unfavorable behaviors can be made the subject of selective breeding, not only maze learning but spontaneous activity (wheel-running), ‘emotionality,’ sound-induced seizures, saccharin preferences, aggressiveness, mating speed, and so on.
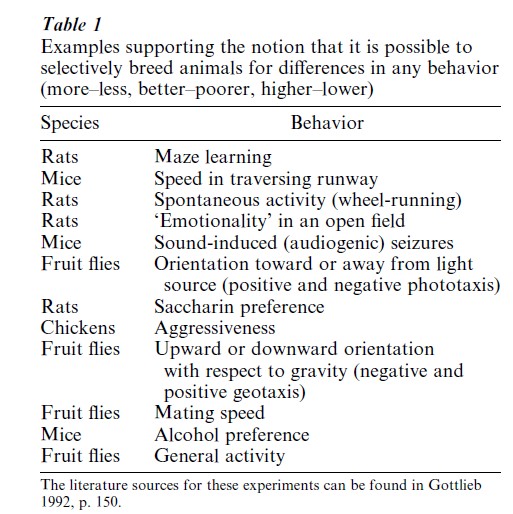
As mentioned above, since all behavioral activities involve both genetic and environmental contributions, it is essential when doing selective breeding experiments to keep the rearing environment the same from generation to generation, otherwise the selective breeding experiments will be a failure. For example, in an experiment by Hood and Cairns (1989), mice were selectively bred for high and low aggressiveness. The standard rearing procedure was to keep the animals in isolation after they were weaned. After the high and low aggressive lines were established, to show the contribution of the isolation rearing procedure to the results of selective breeding Hood and Cairns reared half the animals from each line in social groups rather than in isolation. As can be seen in the right-hand side of Fig. 1, the high aggressive line did not differ from the low aggressive line when the animals were reared in social groups; the high aggressive line was only highly aggressive when reared in isolation (left-hand side of Fig. 1), the rearing condition under which the original selective breeding took place. These results lead into the topic of Sect. 3.
3. Reaction Range vs. Norm Of Reaction
Until very recently some biologists and psychologists held the incorrect belief that genes strictly canalize behavioral development, creating a predictable reaction range when rearing environments are changed from the normal or usual (species-typical) to highly favorable (enriched) or unfavorable (deprived) conditions.
This point of view was expressed by one authority as follows:
Canalization is the genetic limitation of phenotypic development to a few possible phenotypes … The genetic restriction of possible phenotypes is a result of coadapted gene complexes (sets of genes that have evolved together) that buffer the developmental pattern against deviation outside of the normal, species range of variation … The limitation of possible phenotypes to a few rather than many is a fact of genotypes’ ranges of reaction (Scarr-Salapatek 1976, p. 63).
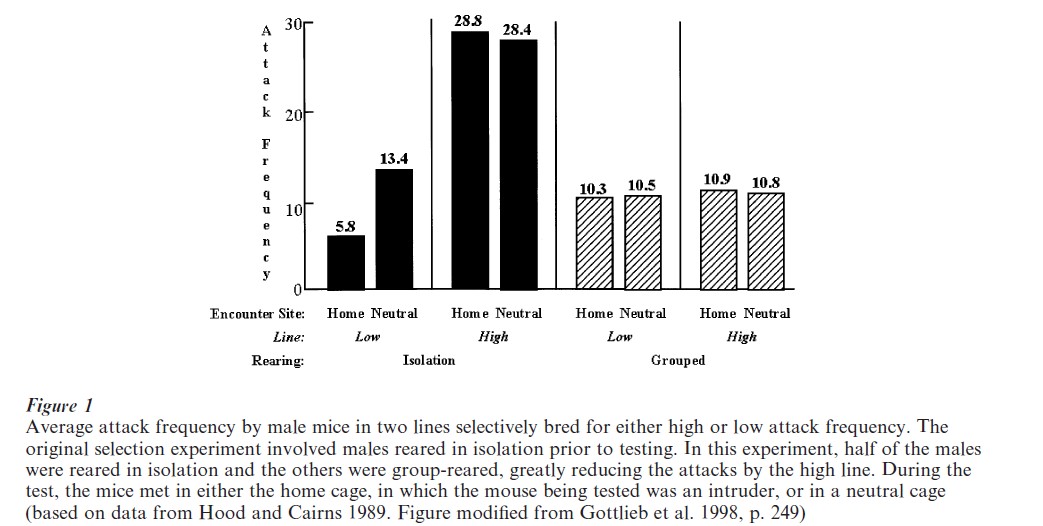
The belief in the sheerly genetic canalization of behavioral development leads quite logically to the reaction-range idea. Thus, the basic idea of a reaction range is that genes determine (canalize) the upper and lower limits of psychological development so that, given a knowledge of the outcome of development under one rearing condition, one can make linear predictions of the upper and lower limits of outcomes under other rearing conditions (see as an example Fig. 2).
In contrast to the reaction range, the norm of reaction holds that the outcome of development is an emergent (epigenetic) property that is a nonlinear result of co-actions occurring at all levels of the developmental system (including but not restricted to genes and environments, as elaborated in Sect. 4). Thus, it is not possible to predict outcomes from one rearing circumstance to the next. Unfortunately, it is the reaction range, not the reaction norm, that has had, until very recently, wide currency in psychology, as evidenced in many textbooks on developmental psychology. These texts reproduced a version of Fig. 2 depicting only the reaction-range concept for learning ability or mean IQ for three hypothesized genotypes reared in very different environments, and not the norm of reaction (far right).
The difference between the predictions of the reaction-range and reaction-norm concepts is testable. As indicated in Fig. 2, the reaction-range idea predicts that the ranking of the different genotypes on IQ or learning ability will remain the same when they are reared in different environments. In the enriched environment, each genotype will show an improvement but the ‘brightest’ genotype will still rank above the other genotypes. In the deprived rearing environment each genotype’s performance will be poorer but the brightest genotype will still rank above the other genotypes. As shown in the far right of Fig. 2, the norm of reaction makes no prediction on relative ranking when the genotypes are reared in environments (enriched or deprived) that differ from the rearing environment in which the selective breeding took place.
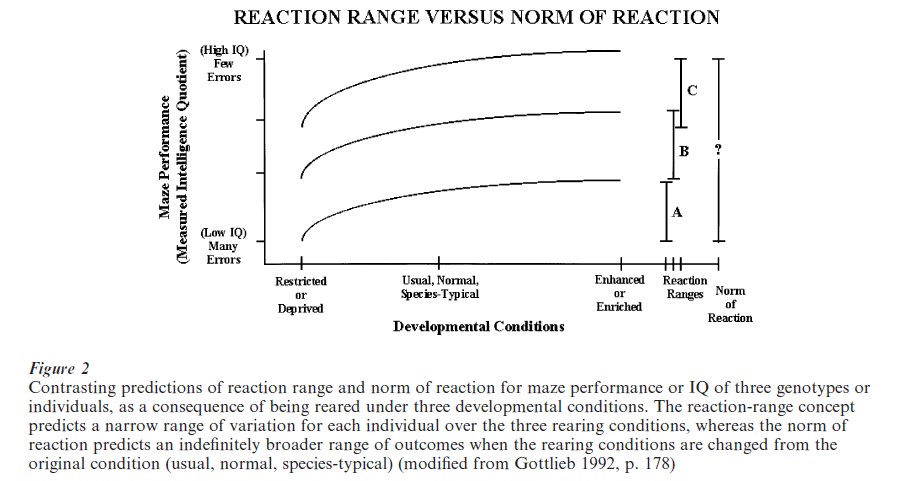
There is only one empirical study in the psychological literature that explicitly addresses the reactionnorm concept, a study by Cooper and Zubek (1958). The results very clearly support the reaction-norm concept. It is interesting to note that the study was carried out with the idea of a reaction range in mind, and it is sometimes erroneously cited as supporting the reaction-range concept. Cooper and Zubek reared maze-bright and maze-dull rats in either an enriched or a restricted environment and then tested them in a Hebb–Williams learning maze. Since they had the reaction-range concept in mind in performing the experiment, Cooper and Zubek thought that the learning of both the bright and dull rats would improve relative to each other under the enriched rearing circumstances and would be poorer relative to each other when reared under the restricted (deprived) condition. This prediction is illustrated in the left-hand side of Fig. 3. Instead, as shown on the right-hand side of Fig. 3, they found equality of performance under both of the new rearing conditions. The dull rats made as few errors as the bright rats after enriched rearing and the bright rats made as many errors as the dull rats after restricted rearing.
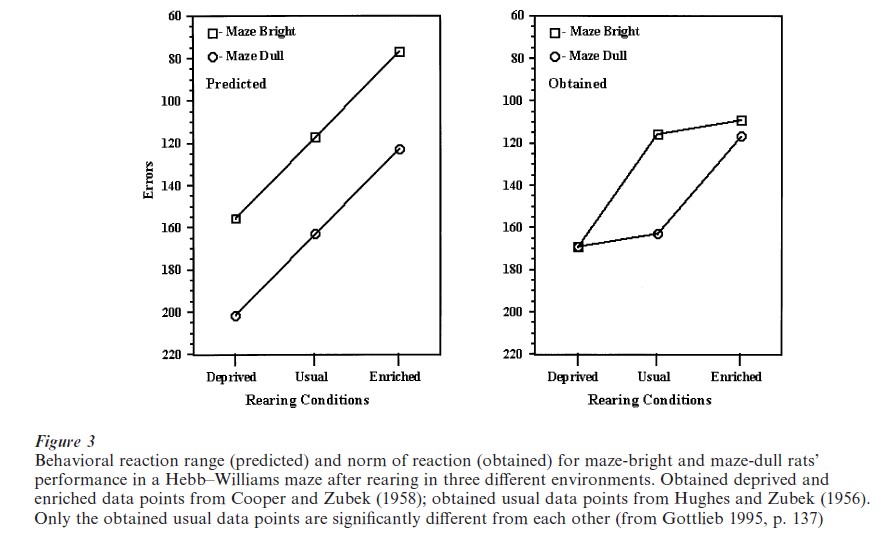
When the so-called bright and dull rats were tested in the Hebb–Williams maze after being reared in their usual way (neither enriched nor deprived), a significant difference between the lines appeared (middle points, right-hand side of Fig. 3). The reason is that this developmental situation repeats the rearing condition under which the original selective breeding for superior and inferior performance was carried out (Hughes and Zubek 1956). If the reaction-range idea were correct and the genes coded for upper and lower limits of learning ability, when these lines were reared under enriched or restricted conditions, the relative differences between them would be preserved. Instead, the experiment shows that the genes are part of a developmental system. The highly specific consequences of rearing under a certain developmental condition were realized by selective breeding under that condition. As called for by the norm-of-reaction concept, selective breeding under one developmental regimen (environment) does not predict outcomes under different rearing conditions. The results of selection depend on the entire developmental manifold, not merely on the genes that are involved.
4. Understanding Development From Genes To Behavior
To provide some insight into what would today constitute a truly developmental behavioral-genetic analysis, Fig. 4 shows a diagram of the four major levels that are involved in moving from genetic activity to environment and back again. This is a fully bidirectional co-actional system, with traditionally trained developmental psychologists working at the Behavior ↔ Environment level, developmental neuropsychologists and neuroscientists at the Neural Activity ↔ Behavior ↔ Environment levels, and biologists beginning now to work on the Genetic Activity ↔ Neural Activity levels. Viewed in this way, developmental understanding or explanation is a multilevel affair involving at least culture, society, immediate social and physical environments, anatomy, physiology, hormones, cytoplasm, and genes. Hierarchical, multilevel, or developmental systems’ analysis is methodologically not theoretically reductionistic in the sense that psychological understanding or explanation does not come from lower levels (or reside in the lower levels), but that both higher and lower levels of analysis are necessary to explain developmental outcomes. The term co-action is used to emphasize the multilevel systems’ nature of developmental analysis.
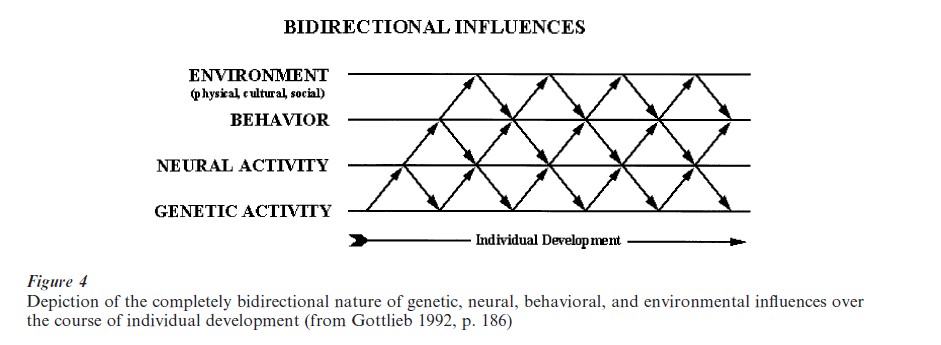
The notion that genetic activity is necessarily influenced by supragenetic factors (factors above the genetic level) may be unfamiliar, in part because of what has come to be known as the Central Dogma of Molecular Biology. The Central Dogma holds that DNA → RNA → Protein, in which DNA serves as a template for RNA which then serves as a template for the synthesis of a protein. In the Central Dogma there are master genes which contain a blueprint of the organism, so ‘information’ is said to flow outward from these master genes providing instructions for individual development. The actual story is rather different. Rather than being encapsulated (or ‘safely sequestered in the nucleus of the cell,’ as some biologists have written), the genes require signals from the internal and external environment to function appropriately. As mentioned above, genes are part of the developmental system of influences; they do not stand outside or apart from the developmental system as the Central Dogma would seem to indicate. Gene expression is influenced by cytoplasmic factors, which provide the immediate environment of the nucleus of the cell. (Structural DNA is found in the nucleus of each cell.) Cytoplasm can be influenced by the external environment of the organism (reviewed by Ho 1984) via the behavior of the organism, which gets the organism into and out of exposure to different environmental influences (physical, social, cultural). Likewise, hormones make their way into the nucleus of the cell and trigger gene expression, and hormones themselves are responsive to the organism’s experiences in the external world. In fact, the action of some genes is dependent on sensory stimulation early in development (reviewed by Arenander and Herschman 1993). Finally, different proteins are formed depending on the particular factors influencing gene expression (Davidson 1986). Hence, genes do not stand outside the developmental system acting as independent causes. They are part of the developmental system (for a detailed discussion, see Wahlsten in press). One of the main reasons for the failure of gene therapy as currently conceptualized and practiced (Rosenberg and Schechter 2000) is that the genes do not function as independent causes; they do not stand outside the developmental system.
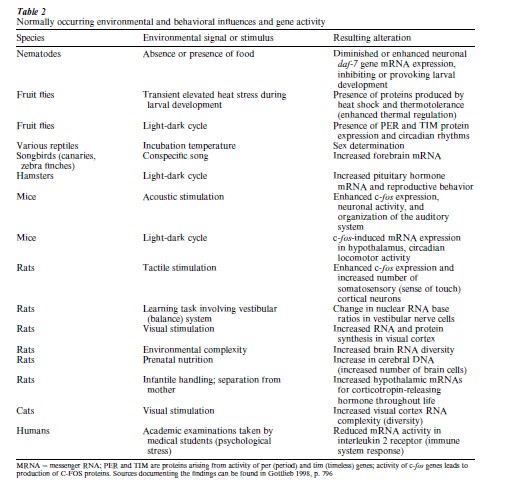
Since there are in fact no master genes telling other genes what to do, in order for genes to function appropriately they require signals from the internal and external environment. Table 2 presents a sampling of the normally occurring external environmental and behavioral signals that activate genes during the normal course of development in a variety of species including humans. The findings summarized in Table 2 substantiate the main point of this research paper: genes and environments necessarily cooperate during the course of individual development.
Bibliography:
- Arenander A T, Herschman H R 1993 Primary response gene expression in the nervous system. In: Loughlin S F, Fallon J H (eds.) Neurotrophic Factors. Academic Press, San Diego, CA
- Cooper R M, Zubek J P 1958 Effects of enriched and restricted early environments on the learning ability of bright and dull rats. Canadian Journal of Psychology 12: 159–64
- Davidson E H 1986 Gene Activity in Early Development. Academic Press, Orlando, FL
- Gottlieb G 1992 Individual Development and Evolution: The Genesis of Novel Behavior. Oxford University Press, New York
- Gottlieb G 1998 Normally occurring environmental and behavioral influences on gene activity: From central dogma to probabilistic epigenesis. Psychological Review 105: 792–802
- Gottlieb G, Wahlsten D, Lickliter R 1998 The significance of biology for human development: A developmental psychobiological systems perspective. In: Damon W, Lerner R M (eds.) Handbook of Child Psychology: Theoretical Models of Human Development, 5th edn. Wiley, New York, Vol. 1
- Ho M-W 1984 Environment and heredity in development and evolution. In: Ho M-W, Saunders P T (eds.) Beyond NeoDarwinism: An Introduction to the New Evolutionary Paradigm. Academic Press, London
- Hood K E, Cairns R B 1989 A developmental-genetic analysis of aggressive behavior in mice: IV. Genotype-environment interaction. Aggressive Behavior 15: 361–80
- Hughes K R, Zubek J P 1956 Effect of glutamic acid on the learning ability of bright and dull rats. I. Administration during infancy. Canadian Journal of Psychology 10: 132–8
- Rosenberg L E, Schechter A N 2000 Gene therapist, heal thyself. Science 287: 1751
- Searle L V 1949 The organization of hereditary maze-brightness and maze-dullness. Genetic Social and General Psychology Monographs 39: 279–325
- Scarr-Salapatek S 1976 Genetic determinants of infant development: An overstated case. In: Lipsett L (ed.) Developmental Psychobiology: The Significance of Infancy. Erlbaum, Hillsdale, NJ
- Tolman E C 1924 The inheritance of maze-learning ability in rats. Journal of Comparative Psychology 4: 1–18
- Tryon R C 1942 Individual diff In: Moss F A (ed.) Comparative Psychology. Prentice-Hall, New York
- Wahlsten D in press Genetics and the development of brain and behavior. In: Valsiner J, Connolly K (eds.) Handbook of Developmental Psychology. Sage, London
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality