
View sample motivational systems research paper. Browse research paper examples for more inspiration. If you need a psychology research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our writing service for professional assistance. We offer high-quality assignments for reasonable rates.
One of the cardinal features of behavior is that it is goaldirected. Animals seek food or water; they avoid predators and explore novel environments. Humans work overtime to buy a new home, get up early to go to the gym, or put their health and happiness in jeopardy to get drugs. The study of motivation attempts to explain why such behaviors occur. From early in the twentieth century when the term motivation was first popularized, scholars have consistently described both activation and direction as the key components of motivated states (e.g., Bindra, 1969; Lashley, 1938; Woodworth, 1918). This notion suggests that in order to understand the antecedent conditions that elicit behavior, one must be able to explain how behavior is stimulated or initiated (i.e., activation) and the reason behavior takes one particular form over another (i.e., direction).
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
The difficulty inherent in the study of such questions is obvious: Hunger and craving are internal states and thus are not directly observable or measurable. Early in the twentieth century, when psychology was struggling to gain acceptance as a science, J. B. Watson (Watson, 1913, 1919) and later B. F. Skinner (Skinner, 1950) convincingly argued that psychology must restrict itself to the study of observable, quantifiable phenomena in order to survive. For psychology this subject matter was behavior. Skinner suggested that the science of behavior would exclude “anything of an observed fact which appeals to events taking place somewhere else, at some other level of observation, described in different terms, and measured, if at all, in different dimensions” (1950, p. 193). In other words, he argued that anything falling within the mental or neurobiological realms—and therefore unobservable—should not be studied within psychology.
This argument had a profound effect on the study of motivation. As motivation is essentially an unobservable phenomenon, many in the field eschewed its study in favor of more observable and describable processes. Instead, research focused on reward and reinforcement. As illustrated in Figure 14.1, motivation is an antecedent condition that instigates behavior. In contrast, reinforcement is a consequence of interaction with the goal object that presumably affects learning and subsequent motivation. In short, reinforcement affects learning, whereas motivation affects the performance of learned behaviors. Unfortunately, inquiry into the neural mechanisms involved in the production of goal-directed behavior has predominantly ignored this distinction; the primary reason was that reinforcement can be operationally defined and measured. Thus, amount of reinforcement (e.g., number of food pellets) and consequent changes in behavior (e.g., number of lever presses or time required to traverse an alley) can be quantified. For this reason, Skinner (and those who followed in his tradition) argued that motivationally significant stimuli should be treated strictly as behavioral reinforcers (Skinner, 1959).
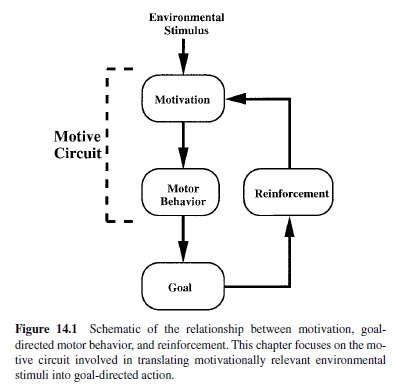
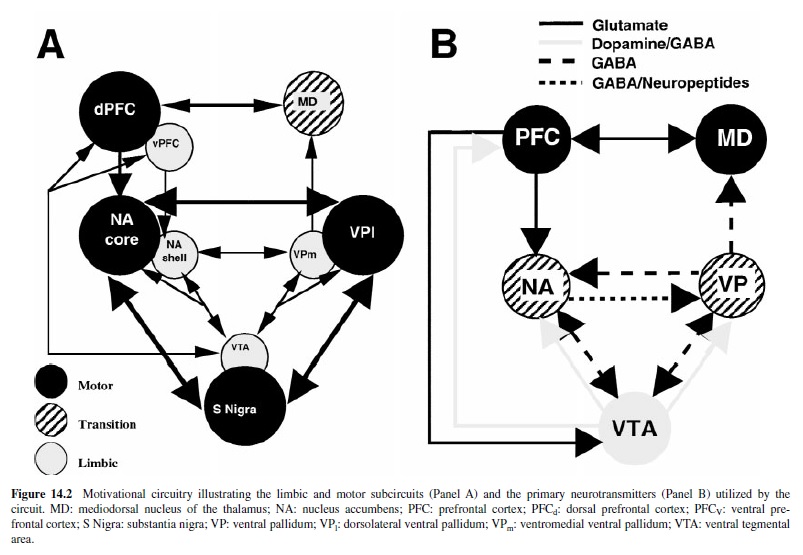
The goal of the present research paper is to review the behavioral, anatomical, and neurobiological evidence regarding the neural substrates of motivated behavior. This involves examination of a motive circuit within the basal forebrain. The motive circuit consists of two parallel circuits that play importantroles in the production of motivation and goal-directed behavior—one predominantly associated with motor function and the other primarily associated with limbic functions (see Figure 14.2, PanelA). It is hypothesized that the motor circuit is critical for the production of well-learned behavioral responses, whereas the limbic circuit is important for processing environmental stimuli and transmitting this information to portions of the motor circuit, thus instigating novel and practiced adaptive motor responses. Within this circuit, dopamine, glutamate, gamma-aminobutyric acid (GABA), and neuropeptides are neurotransmitters (see Figure 14.2, Panel B), each conveying potentially distinct information affecting motivation.The present review is organized around the neruotransmitters in this circuit and in reference to the following hypotheses.
- Glutamate stimulates behavior and the anatomical origin of the activated glutamatergic afferents provides motor memory to provoke the appropriate behavioral response. In addition, under the appropriate conditions glutamate transmission promotes neuroplasticity permitting learning and behavioral adaptations to occur.
- Dopamine supports plasticity and learning by engaging the appropriate cellular machinery to modify neuronal communication, especially excitatory trasmission.
- GABA regulates overall circuit tone and thereby serves to bind or sustain an animal’s motivational state until the goal object can be achieved.
- Neuropeptides contribute to subjective valence.
Thus,the neuroanatomical and neurochemical organization of the motive circuit provides the neural basis of motivation and reinforcement and functions to elicit adaptive motor responses in the presence of motivationally significant stimuli.
Anatomy of the Motive Circuit
Historical Perspectives
Early biological studies of motivated behavior focused on the importance of a few individual nuclei in the production of adaptive motor responses. In this respect, the amygdala garnered early attention—beginning with demonstration that bilateral ablation of temporal lobe, including the amygdala, resulted in dramatic changes in emotionality. Formerly aggressive monkeys became tame and willing to approach normally fear-inducing objects (Klüver & Bucy, 1939). Subsequently, animals with amygdala lesions were shown to have impaired avoidance responding to stimuli that signaled shock (Weiskrantz, 1956) as well as impaired visual discrimination for food reward (B. Jones & Mishkin, 1972). Thus, amygdaloid lesions seemed to produce a deficit in the ability to identify and respond appropriately to biologically significant stimuli. These studies, when combined with anatomical studies demonstrating the amygdala’s connectivity with sensory, autonomic, and motor structures (e.g., Aggleton, Burton, & Passingham, 1980; Herzog & Van Hoesen, 1976; Nauta, 1961) suggested a critical modulatory function in regulating motivated behavior.
The other brain region most frequently implicated in the control of motivated behavior was the nucleus accumbens (NA). In 1954, Olds and Milner discovered that animals would work for electrical simulation of the medial forebrain bundle (Olds & Milner, 1954). Later demonstrations that this effect was largely due to stimulation of dopaminergic afferents to the NA (e.g., Corbett & Wise, 1980; Fibiger & Phillips, 1986)— coupled with emerging evidence that the neurochemical mode of action of many drugs of abuse depended on the accumbens (e.g., Kornetsky & Espositio, 1979; Wise, 1982)—indicated a central role in goal-directed behavior. Behavioral evidence, considered in conjunction with the connectivity of the NA with limbic and cortical structures, suggested a probable role in behavior elicited by at least positive motivational stimuli.
Whereas research on the functions of the amygdala arose from a tradition interested in examining its emotional and motivational properties, research on the NA arose from a tradition primarily interested in identifying the neural substrates of reinforcement and reward, an endeavor that has only recently begun to implicate the amygdala. Thus, one perspective valued motivation as a concept, whereas the other was more Skinnerian in orientation. In the now-classic formulation, Mogenson, Jones, and Yim (1980) moved beyond both the separate ideologies and the single nucleus approach in order to suggest that the amygdala and NA form part of a circuit that functions to integrate limbic information and elicit appropriate behavioral responses. This circuit—termed herein the motive circuit—was presumed to integrate signals from limbic structures about motivationally important stimuli and recruit motor structures to elicit adaptive behavioral output (i.e., translate motivation into action). More recently, Heimer, Alheid, and Zahm (1993) formalized the anatomical interrelationships between the amygdala and NA in motivation as the extended amygdala. This interconnected series of nuclei—including the central nucleus of the amygdala, bed nucleus of the stria terminalis, medial ventral pallidum (VP), and ventromedial NA—is hypothesized to be a prime contributor of emotional context.
Subcortical Circuitry
Figure 14.2 shows a schematic picture of the circuitry that has been implicated in translating motivational stimuli into the production of goal-directed behavior. A central component of the motive circuit is a trio of interconnected nuclei that display tight topographical organization such that they form two parallel loops through the ventral mesencephalon, ventral striatum, and ventral pallidum (see Figure 14.2, Panel A). The mesoaccumbens system consists of a welldocumented dopaminergic projection from the ventral tegmental area (VTA) to the nucleus accumbens in the ventral striatum (Beckstead, Domesick, & Nauta, 1979; Fallon & Moore, 1978), although up to 20% of the pathway contains GABA (Carr & Sesack, 2000a). This projection is reciprocal and topographically organized such that the VTA innervates and receives innervation from ventromedial portions of the nucleus accumbens, termed the shell (NAs; Heimer, Zahm, Churchill, Kalivas, & Wohltmann, 1991; Swanson, 1982). Accumbal projections to the VTAcontain GABA, dynorphin, and substance P (Churchill, Dilts, & Kalivas, 1990; Napier, Mitrovic, Churchill, Klitenick, & Kalivas, 1995). While the VTA also innervates the medial portions of the accumbens, termed the core (NAc; Heimer et al., 1991), reciprocal projections with the NAc arise from the substantia nigra (SN), which is classically considered a component of the extrapyramidal motor system (Heimer et al., 1991).
The topography of the loop is maintained in the efferents from the NA to the VP. Thus, the NAs projects to ventromedial portions of the VP (VPm), while the NAc projects primarily to the dorsolateral, subcomissural VP (VPl; Zahm & Heimer, 1990). The striatopallidal projections have been shown to contain GABA, enkephalin, substance P, and neurotensin (NT; Churchill et al., 1990; Napier et al., 1995), whereas reciprocal projections seem to be primarily GABAergic (Churchill & Kalivas, 1994). Like the NA, the VP exhibits medioloateral topography in its innervation of the mesencephalon, with the VPm providing GABAergic innervation of the VTA and the VPl innervating the SN (Kalivas, Churchill, & Klitenick, 1993b; Zahm & Heimer, 1990). However, reciprocal innervation of the VP is not as discrete. The VTA projects to both the VPm and the VPl, while the SN shows little if any innervation of the VP (Klitenick, Deutch, Churchill, & Kalivas, 1992).
Prefrontal Cortical Input
Afferent innervation of the subcortical circuit arises from a number of sources. Primary among these is the medial prefrontal cortex (PFC) because it maintains topographic connectivity with both the VTA and the NA, and thus forms an extension of the parallel subcortical circuitry. Both the dorsal (PFCd) and ventral (PFCv) prefrontal cortices receive mesocortical dopamine projections from the VTA (Füxe, Hökfelt, Johannson, Lidbrink, & Ljungdahl, 1974), which—like the mesoaccumbens system—has a significant (up to 40%) GABAergic component (Carr & Sesack, 2000a). Additionally, both the PFCd and the PFCv send reciprocal projections back to the VTA, and the PFCd also sends a projection to SN (Sesack, Deutch, Roth, & Bunney, 1989). Whereas the PFCv and PFCd show some degree of mediolateral topography in corticofugal innervation of the ventral mesencephalon, the PFC displays very discrete target specificity in its innervation of the NA. The dorsal PFCd projects selectively to the NAc, and the PFCv projects to the NAs (Berendse, Galis-de Graaf, & Groenewegen, 1992; Sesack et al., 1989). The main transmitter of the efferent PFC projections is glutamate (e.g., Christie, Summers, Stephenson, Cook, & Beart, 1987; Fonnum, Storm-Mathiasen, & Divac, 1981).
Allocortical Afferents
In addition to excitatory afferents arising from the PFC are allocorticalglutamatergicinputs,includingthosefromthebasolateral amygdala (BLA) and the hippocampus (Figure 14.3). The BLA sends excitatory glutamatergic projections to many nuclei within the motive circuit, including the NA (Wright, Beijer, & Groenewegen, 1996), medial prefrontal cortex (mPFC; Bacon, Headlam, Gabbott, & Smith, 1996; McDonald, 1996), VTA (Otake & Nakamura, 2000), VP (Grove, 1988) and mediodorsal thalamus (MD; Reardon & Mitrofanis, 2000). In turn, the BLA receives a reciprocal dopaminergic projection from the VTA (Brinley-Reed & McDonald, 1999), as well as afferents from the mPFC (Groenewegen, Berendse, Wolters, & Lohman, 1990; McDonald, Mascagni, & Guo, 1996) and VP (Groenewegen, Berendse, & Haber, 1993). Notably, the BLA is also densely innervated by thalamus and sensory cortex (Groenewegen et al., 1990), suggesting that it integrates sensory information about reinforcing stimuli and can then relay this information to the motive circuit.
In addition to the BLA, it is also important to consider the innvervation of the VTAand substantia nigra from the central nucleus of the amygdala (Haber & Fudge, 2000). This is thought to be a primary source of enkephalin and neurotensin innervation to the VTA. Likewise, the VTA has a substantive dopaminergic and GABAergic projection to the central nucleus. As part of the extended amygdala (discussed later in this research paper), the central nucleus also provides innervation to the VPm and NAs (Heimer et al., 1993). Precisely how the BLA and central nucleus integrate motivationally relevant stimuli and promote learning is a subject of much recent experimental scrutiny; the reader is referred to excellent articles on this topic (Hatfield, Han, Gallagher, & Holland, 1996; Hitchcott & Phillips, 1998; Parkinson, Robbins, & Everitt, 2000). However, it is noteworthy for purposes of this review that the central nucleus receives dense excitatory input from the BLA and not only acts as a conduit for providing peptidergic modulation of some nuclei in the motive circuit, but also provides adaptive modulation of autonomic responses to motivationally relevant stimuli.
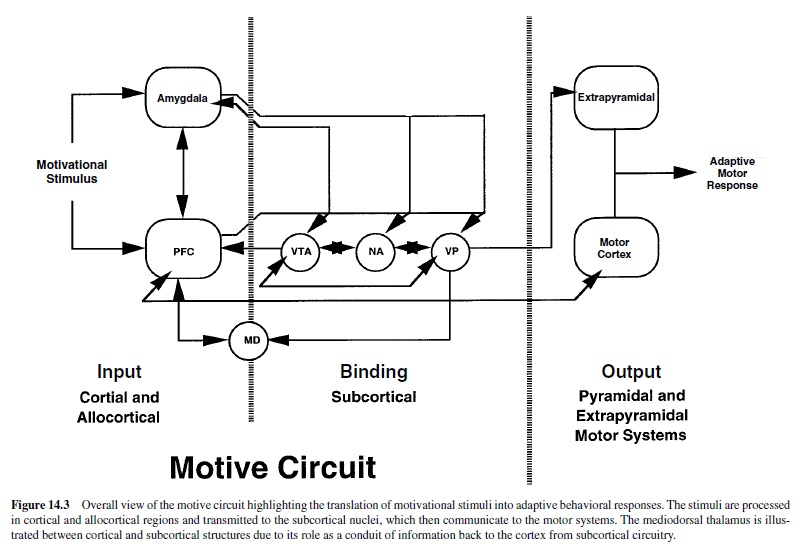
The hippocampus—primarily via the ventral subiculum— sends glutamatergic efferents to the NA (Groenewegen, Vermeulen-Van der Zee, Te Kortschot, & Witter, 1987; Kelley & Domesick, 1982) and PFC (Carr & Sesack, 1996), but not to the VP (Yang & Mogenson, 1985). It receives a dopaminergic projection from the VTA (Gasbarri, Verney, Innocenzi, Campana, & Pacitti, 1994).
Motor and Limbic Subcircuits
An inspection of Figure 14.2 (A) and the preceding description reveals that there are two subcircuits that comprise the larger motive circuit—one comprosed of predominantly limbic structures and one of primarily of motor structures. Thus, the VTA, NAs, and VPm are associated with limbic structures like the PFCv, BLA, hippocampus, and bed nucleus of the stria terminalis (BNST). In fact, recent conceptualizations of the ventral forebrain suggest that the centromedial amygdala, sublenticular VP, BNST, and NAs form a continuous network termed the extended amygdala (Kalivas, Churchill, & Klitenick, 1993a). Conversely, the PFCd, NAc, VPl form contacts with motor structures like the SN, motor cortex, pedunculopontine nucleus (PPN) and subthalamic nucleus. Thus, there are two relatively closed loop systems that can integrate motor and limbic information separately (see Kalivas et al., 1993a, for more details).
Of course, for an individual to effectively integrate incoming motivational stimuli and emit appropriate behavioral responses, there must be interplay between the motor and limbic systems. There are two major pathways that permit such interaction. The first is via the VTA that forms reciprocal connections with both the PFCd and the PFCv, as well as both the NAc and NAs. Additionally, although it receives a projection only from the VPm, it sends projections to both the VPm and VPl. Thus the permissive topography of VTA efferent projections within the motive circuit positions it to influence the activity of both the motor and limbic subcircuits. The second means of interaction between these two systems is the MD. The VP sends a prominent GABAergic efferent to the MD, with the primary contribution coming from the VPm and only minor involvement of the VPl (Mogenson, Ciriello, Garland, & Wu, 1987; Zahm, Williams, & Wohltman, 1996). The MD does not send a reciprocal projection to the VP, but there is reciprocal glutamatergic innervation of the PFCd (Groenewegen, 1988; Kuroda, Murakami, Shinkai, Ojima, & Kishi, 1995). Thus, the MD receives information from the limbic circuit via the VPm but sends a projection to the motor associated PFCd, consequently forming a bridge between limbic and motor circuitry.
It is noteworthy that communication between the limbic and motor subcircuits is rectified to permit more direct information flow from limbic to motor, whereas flow in the reverse direction requires multisynaptic communication. Three rectified projections service this preferential flow from limbic to motor subcircuits, including (a) the dopaminergic and GABAergic innervation by the VTA of the NAc and VPm, (b) the GABAergic projection from the VPm to the MD, and (c) the glutamatergic projection from the PFC to the NA. The location of rectified information flow is strategic for the movement of information from limbic to motor subcircuit, and it has been suggested that information may spiral outwards from the more medial limbic nuclei to the more lateral and dorsal motor nuclei in the motive circuit (Haber & Fudge, 1997; Zahm & Brog, 1992).
Motivationally relevant information can exit the motive circuit to the motor system via a number of different pathways. Hence, the VPl projects to the PPN, subthalamic nucleus, SN, and subsequently to all parts of the extrapyramidal motor system (Haber, Groenewegen, Grove, & Nauta, 1985). There is also a projection to motor cortex that arises from the PFCd (Zahm & Brog, 1992). Finally, the SN receives a projection directly from the NAc. Thus, the motive circuit has several conduits by which it can influence motor behavior following presentation of motivationally relevant stimuli.
Dopaminergic Effects on Motivated Behavior
There is abundant evidence that midbrain dopamine systems play an important role in motivated behavior (for reviews, see Ettenberg, 1989; Koob, 1992; Wise, 1982). Thus, administration of dopamine receptor antagonist drugs has been shown to disrupt responding for a variety of reinforcers, including food and water (e.g., Ettenberg & Horvitz, 1990; Gerber, Sing, & Wise, 1981), electrical brain stimulation (e.g., Fouiezos, Hansson, &Wise, 1978; Stellar, Kelley, & Corbett, 1983), and drugs of abuse (e.g., Bozarth & Wise, 1981; de Wit & Wise, 1977;Yokel&Wise,1976).Despitethewell-documentedrole of dopamine (DA) in regulating goal-directed behavior, the specific function that DA serves is emerging, but remains uncertain. Theories suggest a role for DA in everything from reward (e.g., Schultz, 1998; Wise & Rompre, 1989) to response initiation or selection (Beninger, 1983; Salamone, Kurth, McCullough, Sokolowski, & Cousins, 1993) to motivation-wanting (Robinson & Berridge, 1993). The following is an attempt to integrate what is known about dopaminergic function in order to frame its role in the production of motivated behavior.
Many postulates suggest that midbrain DA neurons function in reward, indicating that they govern behavior directed toward appetitive (rather than aversive) stimuli. Such suggestions seem at best incomplete because DA neurons have been shown to respond to presentation of aversive as well as appetitive stimuli (e.g., Doherty & Gratton, 1992; Louilot, Le Moal, & Simon, 1986). Additionally, DAreceptor antagonism has been shown to disrupt learning about aversive stimuli (Salamone, 1994). Furthermore, DA neurons do not fire in a temporal pattern consistent with a role in pleasure or hedonics. Thus, after reward is expected, DAneurons have been shown to respond not to presentation of the reward itself but instead to presentation of a stimulus that is most predictive of the reward—even before it is presented (Schultz, Apicella, & Ljungberg, 1993). For these reasons, theories of DA function that depend on notions of hedonics and reward have largely been dismissed. However, the notion that DA serves to increase the frequency of future behavior directed toward a stimulus suggests in essence that DA functions in appetitive learning situations is a topic to which we return later in this research paper.
Dopamine Mediates the Learning of Motivational Responding but Not the Emission of Motivated Behavior
Purported roles for DAin wanting or craving, as well as in response initiation or response selection, are versions of motivational theories of DAergic function and are very influential in contemporary thinking about motivated behavior. They suggest that DA is involved in the energizing or directing of behavior toward the appropriate goal. However, behavioral evidence suggests that DA receptor antagonism leaves motivational processes very much intact. For example, animals can be trained to run a straight alley when presented with an olfactory cue (S+ ) predictive of either food or drug reinforcement in the goal box. Following DA receptor antagonist treatment, such animals still traverse the alley normally when presented with the reinforcement-predictive cue (McFarland & Ettenberg, 1995, 1998). Furthermore, in subjects having undergone training to run an alley for heroin reinforcement and a subsequent period of extinction (with no cues or reinforcement available), haloperidol does not block the ability of the S + to reinstate drug-seeking behavior (McFarland & Ettenberg, 1997). Additionally, the ability of an S+ conditioned in this fashion to elicit conditioned locomotor activation and a conditioned place preference remain intact during dopamine receptor antagonist treatment (McFarland & Ettenberg, 1999). Together these data strongly suggest that the motivational capacity of the S+ stimulus (i.e., its ability to activate and direct behavior) remains intact despite DA receptor blockade.
Studies examining the role of conditioned stimuli in behavioral activation have produced comparable results. Horvitz and Ettenberg (1991) showed that administration of pimozide did not reduce locomotor activity in the presence of a stimulus previously paired with food delivery. This suggests that the motivational properties of food-paired stimuli are left intact. Such data are also consistent with demonstrations that environments or stimuli previously paired with amphetamine reinforcement retained their conditioned behavior-activating effects under dopamine receptor antagonist challenge (Beninger & Hahn, 1983; Beninger & Herz, 1986).Additionally, preferential responding on a lever associated with conditioned reinforcement is preserved following dopaminergic denervation of the ventral striatum (Everitt & Robbins, 1992; Robbins, Cador, Taylor, & Everitt, 1989). Thus it seems that the motivating capacity of reinforcementassociated cues remains intact following disruption of DA function.
When subjects are actively engaged in operant responding, administration of a DA receptor antagonist produces one of two behavioral patterns. Low doses produce increases in responding similar to those seen when the reinforcer is diminished (e.g., Ettenberg, Pettit, Bloom, & Koob, 1982; Schneider, Davis, Watson, & Smith, 1990). High doses produce withinsession declines in operant behavior, similar to extinction curves that result from removal of the reinforcer (e.g., Gallistel, Boytim, Gomita, & Klebanoff, 1982; Gerber, Sing, & Wise, 1981; Wise, 1978). The fact that in both situations, animals will initiate responding and do so with normal (or near-normal) response latencies suggests that the motivation of these subjects toengageingoal-orientedbehaviorisverymuchintact.
Franklin and McCoy (1979) trained animals to press a lever in order to receive electrical brain stimulation. They demonstrated that when pretreated with pimozide, animals showed an extinction-like pattern of responding. However, presentation of a conditioned stimulus (CS) that was previously paired with brain stimulation reward successfully reinstated operant responding. Thus, subjects maintained motivational responding to a reward-paired stimulus despite the reinforcement decrement that presumably led to the progressive decline in responding through the initial course of the session. Similarly, Gallistel et al. (1982) showed that although dopamine antagonists elevated brain reward thresholds for intracranial stimulation in a runway paradigm, they did not prevent the motivational effects of priming stimulation that incited animals to run the alley in the first place. Taken together, these data suggest that DA receptor antagonism—although it is capable of blocking the ability of reinforcing stimuli to maintain responding—does not alter the motivation to seek reinforcement.
Further evidence that motivational processes remain intact during DA receptor antagonism comes from choice experiments. In such experiments, subjects are allowed to choose between two alternative responses: one that leads to reinforcer delivery and one that does not. Doses of dopamine receptor antagonist drugs that are sufficient to disrupt operant response rates have little effect on response choices in lever-press (Bowers, Hamilton, Zacharo, & Anisman, 1985; Evenden & Robbins, 1983) or T-maze (Tombaugh, Szostack, & Mills, 1983) tasks. Rats still prefer to make a response that has previously led to reinforcement over one that has not, even following challenge with DA receptor antagonists. Taken together, the data described suggest that the fundamental aspects of motivation remain intact despite disruption of DA transmission.
Although the midbrain DAsystem does not seem to signal either reward or motivation, it is clear that intact DAergic function is important for both the acquisition and maintenance of operant responding (for reviews, see Beninger, 1983; di Chiara, Acquas, Tanda, & Cadoni, 1993; Kiyatkin, 1995). Thus, DAmust serve a function related to the learning and maintenance of motivated responding, while the emission of previously learned behavior progresses independent of dopamine receptor activation.
Dopamine-Stimulated Plasticity Within Motivational Circuitry
An examination of the firing pattern of DA neurons reveals that most DA neurons display phasic activation after novel stimuli and after delivery of primary reinforcers (e.g., food). Additionally, when a biologically significant stimulus is predicted by an environmental cue, with experience DA neurons come to respond to the predictive cue rather than to the reinforcer itself. Such changes in firing rate produce a pattern of responding whereby DA neurons increase firing to betterthan-expected outcomes, remain unaffected by predictable outcomes, and decrease firing in response to worse-thanexpected outcomes (for a review, see Schultz, 1998). Thus, DA neurons respond to the difference between actual reward and expected reward, not the presence of reward itself. This suggests that the function of DA within the production of goaldirected behavior is to signal the need to create an adaptive behavioral response—that is, to promote neuronal plasticity.
Such a suggestion is consistent with evidence regarding the anatomical location of DAsynapses.As described earlier, the primary dopaminergic projections within the motive circuit are the mesolimbic and mesocortical systems (Beckstead et al., 1979; Fallon & Moore, 1978; Füxe et al., 1974). Thus, dopamine synapses form on medium spiny neurons of the NA(both core and shell) as well as pyramidal neurons of PFC (both dorsal and ventral). Additionally, the BLA receives dopaminergic innervation. Anatomical studies indicate that DA afferents are well situated to modulate or gate the probability of cells being activated (O’Donnell & Grace, 1995). Thus, DAsynapses in both the PFC and NAtend to be located proximal to excitatory contact (Sesack & Pickel, 1990; Yang, Seamans, & Gorelova, 1999)—for example, in the NA, excitatory inputs form on the head of the spine and dopamine terminals synapse on the neck (Carr, O’Donnell, Card, & Sesack, 1999; Smiley & Goldman-Rakic, 1993). Thus, from a purely anatomical perspective, DA synapses seem to be poised to modulate incoming excitatory information.
Ample electrophysiological data also suggest that DA is capable of modulating the efficiency of neuronal responses to other inputs—particularly to glutamate—either supporting or diminishing neuronal activity, depending on the quality of excitatory inputs received by target cells (O’Donnell & Grace, 1995). Both pyramidal cells in the mPFC and spiny cells of the accumbens have been shown to exist in a bistable state (Bazhenov, Timofeev, Steriade, & Sejnowski, 1998; O’Donnell & Grace, 1995; Timofeev, Grenier, & Steriade, 1998;Yim & Mogenson, 1988). Thus, cells fluctuate between a down state in which membrane potential is relatively hyperpolarized and an up state in which membrane potential is relatively depolarized. Dopamine tends to inhibit cells in the down state but tends to excite cells in the up state (HernandezLopez, Bargas, Surmeier, Reyes, & Galarraga,1997; Kiyatkin & Rebec, 1999, O’Donnell, Greene, Pabello, Lewis, & Grace, 1999;Yang & Seamans, 1996). Thus, if there is more depolarizing (i.e., glutamatergic) input to a cell, DAD1 receptor activation increases the duration of depolarization via increasing a calcium conductance (Hernandez-Lopez et al., 1997). In the absence of depolarizing input, DA will support the inactive state via D2 receptor activation of potassium conductances (O’Donnell & Grace, 1996).
Dopamine appears to serve a similar role within the BLA, wheretherearetwotypesofneurons—inhibitoryinterneurons and pyramidal-like projection neurons. Stimulation of DAreceptors in the BLA increases the firing rate of interneurons, thereby decreasing the firing rate of projection neurons. Furthermore, DA attenuates activation of pyramidal cells in the BLA that is elicited by electrical stimulation of the PFC and MD while potentiating the responses evoked by electrical stimulation of sensory association cortex (Rosenkrantz & Grace, 1999). This organization is suggested to produce a global filtration of inputs such that—upon presentation of an affective stimulus—there is a potentiation of the strongest sensory input and a concomitant dampening of cortical inhibition, thereby augmenting the response to affective stimuli. When considered as a whole, DAseems to increase the signalto-noise ratio and consequently gate the flow of information within the motive circuit (Le Moal & Simon, 1991; Rosenkrantz & Grace, 1999).
The pattern of DAinnervation of its target structures is also consistent with a general filtration and modulatory function. Dopaminergic projections to target structures are very divergent, with each axon being highly ramified (Anden, Füxe, Hamberger, & Hökfelt, 1996; Percheron, Francois, Yelnik, & Fenelon, 1989). Nearly every striatal neuron and many cortical neurons receive dopaminergic innervation. Additionally, these neurons display homogeneous and synchronous responsivity following presentation of motivationally significant stimuli that activate DA cells. Thus, DA neurons broadcast a global wave of activity to the NAand PFC, rather than a stimulus- or response-specific signal (Schultz, 1998). Such a pattern of responding is suited to simultaneous modulation of ongoing activity in these forebrain and allocortical structures.
A Role for Dopamine-Induced Plasticity in the Acquisition of Adaptive Behavior
The data previously outlined suggest that behavioral responding to motivationally relevant stimuli proceeds in at least two phases: the acquisition of a response and the maintenance of a response. During the acquisition phase, synaptic DA is increased by presentation of primary reinforcers or novel stimuli. This DA signal can specifically strengthen those synapses receiving simultaneous excitatory glutamatergic input (e.g., corticostriatal or amygdalostriatal). In this fashion, DA would serve to facilitate the learning of adaptive behavioral responses, as well as increase access of limbic and cortical structures to the motor system. With repeated presentations of motivationally relevant stimuli (either primary or conditioned), these same excitatory inputs would be recruited and strengthened such that they no longer require DAergic influence to elicit motor output. Thus, the primary function of DA is to facilitate synaptic (and behavioral) plasticity, rather than to directly elicitat motor responses. This helps to explain why behavioral data show that animals do not acquire behavioral responses when DA transmission is disrupted and nevertheless exhibit previously learned behaviors (discussed previously). This explanation is also consistent with the observation that the activity of DA neurons fails to discriminate among different salient stimuli—regardless of valence—or among different sensory modalities. Thus, DA facilitates the learning of goal-directed responses in general, rather than specific motor responses to specific stimuli.
The involvement of DA in both the acquisition and maintenance of operant responding has been difficult to explain with a single theory of DA function. Theories emphasizing the modulatory effects of DAin learning can explain acquisition effects; however, they typically fail to explain effects on maintenance. Thus, if the inhibition of DAneurotransmission blocks plasticity, it should cause a kind of behavioral and neuronal inflexibility that leads to a decrease in responding for reinforcers and a perseverance in previously learned behavioral patterns. However, if one remembers that both increases and decreases in firing rates of DAergic neurons have functional implications, then a possible explanation presents itself. As discussed earlier, increases in DA firing rates seem able to support behavioral and neuronal plasticity leading to the learning of new adaptive responses. Similarly, depressed DA transmission (like that resulting from DA receptor antagonism) provides a signal indicating a less-than-expected outcome. From a functional perspective, such an error signal could lead to compensatory adaptations that would weaken the strength and persistence of the preceding behavior. Thus, it seems possible that both an augmentation and a diminution in DA cell firing rates would elicit behavioral plasticity resulting in a change in behavioral output.
Glutamate and Motivated Behavior
Evidence regarding the role of glutamate in the production of motivated behavior has been slower to emerge; this is largely due to the ubiquitous distribution of glutamate within the brain and the relative difficulty in interpreting neurochemical changes in glutamate transmission (McGeer, Eccles, & McGeer,1987;Timmerman&Westerink,1997).Thepresence of glutamate in so many regions has made systemic administration of glutamatergic drugs difficult to characterize in terms of site of action. For this reason, many of the behavioral data helping to elucidate the role of glutamate in goal-directed behavior are indirect. Some information comes from the memory literature, in which food reinforcement is used to encourage subjects to engage in memory-related tasks, and glutamate plays a critical function in long-term potentiation (LTP), which serves as a cellular model of learning and memory (Bliss & Collingridge, 1993; Fonnum, Myhrer, Paulsen, Wangen, & Oksengard, 1995). Thus, glutamate activity is required for the cellular changes that are presumed to underlie learning and memory. Additionally, alterations in glutamate function have been shown to produce concomitant changes in goal-directed behavioral indexes of memory function. Glutamate receptor antagonists have been shown to impair LTP and the establishment of spatial memory in mice (Morris, Anderson, Lynch, & Baudry, 1986), whereas drugs that increase glutamate transmission have been shown to improve memory performance in both mice and individuals with schizophrenia (Firth et al., 1995; Nishikawa, Takasima, & Toru, 1983). Glutamate receptor agonists have been shown to increase speed of performance in a radial-arm maze task and to improve retention of spatial information (Davis et al., 1997). Thus, it seems that increased glutamate transmission facilitates—whereas decreased glutamate transmission impairs—a subject’s ability to earn reinforcement in a memory task.
Modulating Glutamate Transmission in the Motive Circuit
The majority of evidence that glutamate acts in motivational circuitry to modulate motivated behavior comes from studies of motor activity. Glutamate receptors are both metabotropic (mGluR) and ionotropic (iGluR), and both classes of receptor have been implicated in the production oflocomotoractivation.InfusionoftheiGluRagonistsAMPA (alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionate) and NMDA (N-methyl-D-aspartate) directly into the NA results in locomotor activation that is DA dependent (e.g., Boldry, Willins, Wallace, & Uretsky, 1991; Donzanti & Uretsky, 1983). Stimulation of iGluRs in the VTA also elicits motor activation, and this effect arises primarily via activating dopamine neurons (Kalivas, Duffy, & Barrow, 1989). Additionally,AMPAandNMDAbothstimulatemotoractivity when infused into the VP (Churchill & Kalivas, 1999; Shreve &Uretsky,1989).Thismotorstimulationisblockedbyinactivation of the midbrain extrapyramidal area, but not by inactivation of the MD (Churchill & Kalivas, 1999), suggesting that glutamate receptor activation within the VP directly activates motor output to the extrapyramidal motor system. Whereas activation of the NA, VTA, or VP with iGluR agonists elicit locomotor behavior, the infusion of AMPA and NMDAreceptor antagonists into the NAor VP decreases the hypermotility elicited by psychomotor stimulant drugs (Willins, Wallace, Miller, & Uretsky, 1992). Furthermore, either pharmacological or electrical stimulation of the ventral subiculum results in hypermotility that is reversed by blockade of ionotropic receptors in the NA (Mogenson & Nielson, 1984; Pornnarin, Floresco, & Phillips, 2000), suggesting that activation of the NA via stimulating glutamatergic afferents also elicits locomotor activation.
Akin to iGluR stimulation, the ability of mGluR agonists to elicit motor activation can be demonstrated in studies showing that intra-accumbens infusion produces increased locomotion when infused bilaterally (Attarian & Almaric, 1997; Kim & Vezina, 1997) and contralateral rotation when infused unilaterally (Kaatz & Albin, 1995; Sacaan, Bymaster, & Schoepp, 1992). Similarly, mGluR activation in the VTAis capable of inducing dopamine-dependent motor activity (Sacaan et al., 1992; Vezina & Kim, 1999), whereas blocking mGluRs in the VTA produced no change in ongoing motor activity (Kim & Vezina, 1998). Together, these studies demonstrate that activating glutamate transmission can elicit motor-activating effects in all subcortical nuclei of the motive circuit, including the VTA, nucleus accumbens, and VP.
Although the behavioral studies directly examining the role of glutamate in the production of goal-directed behavior are sparse relative to those investigating dopamine, recent data suggest it plays a crucial role. Thus, in animals trained to self-administer cocaine, stimulation of ionotropic glutamate receptors within the NA elicits a reinstatement of drugseeking behavior following a period of abstinence (Cornish, Duffy, & Kalivas, 1999). Furthermore, AMPA—but not DA—receptor antagonism within the NA blocks the reinstatement of drug-seeking elicited by cocaine injection (Cornish & Kalivas, 2000). Moreover, blockade of NMDA receptors—but not D2—receptors disrupts the guidance of instrumental behavior in a reaction time task. Thus, in welltrained animals, stimuli predicting larger or more favorable reinforcement elicit faster responding than do stimuli predicting smaller or less preferred reinforcement. Administration of APV (2-amino-5-phosphonoraleric acid, an NMDA receptor antagonist)—but not haloperidol—into the NA blocked this shortening of reaction times, suggesting that glutamate receptor activation was critically involved in eliciting such fast responding (Hauber, Bohn, & Giertler, 2000). Additionally, it has been demonstrated that blockade of AMPAkainate receptors within the NAs—but not the NAc—elicits feeding behavior even in satiated rats (Maldonado-Irizarry, Swanson, & Kelley, 1995), an effect that has been attributed to disruption of a tonic excitatory input to the shell, thereby inhibiting the firing rate of a population of neurons within the NA. Taken together, these behavioral data suggest that glutamate input to the NA plays a critical role in eliciting of goal-directed behaviors, including feeding and drug seeking.
Organization of Glutamatergic Projections Within the Motive Circuit
Some of the best evidence implicating glutamate in the control of motivated behavior is anatomical. Glutamate provides the major excitatory input to the ventral striatum, arising from the PFC, BLA, and hippocampus (for a review, see Parent & Hazrati, 1995). Additionally, neurons within each of these regions respond to presentation of motivationally relevant stimuli. Some BLAneurons have been shown to respond generally to presentation of both positive and negative reinforcers, as well as unfamiliar stimuli, whereas others respond more specifically only to a single reinforcing stimulus. Notably, stimulus-specific neurons show reversible firing rates when the affective value of the stimulus is diminished (e.g., salting a piece of watermelon). Furthermore, activity of neurons within the amygdala does not relate directly to motor output; thus, firing seems to signal ongoing recognition of the affective significance of complex stimuli (for a review, see Ono, Nishijo, & Nishino, 2000).
Firing of pyramidal neurons within the PFC (or orbital prefrontal cortex in primates) has been correlated with presentation of a reinforcer and a stimulus predictive of reinforcer delivery (i.e., reward expectancy; Gray, Maldonado, Wilson, & McNaughton, 1995; Watanabe, 1996). The firing rates of these neurons have been shown to be sensitive to reward preference, stressing the importance of motivational value for responsiveness (Tremblay & Schultz, 1999). Similar to those in the amygdala, neurons in the PFC showed rapid response modifications that were partially reversible following reinforcer devaluation (Rolls, Critchlet, Mason, & Wakeman, 1996).
Processing and anticipation of reward have also been shown to affect firing patterns of hippocampal neurons (Tamura, Ono, Fukuda, & Nishijo, 1992); this has particular relevance for our understanding of motivated behavior because hippocampal—but not cortical, amygdaloid or thalamic—glutamatergic afferents to the nucleus accumbens have the ability to regulate the transition of medium spiny neurons into the up state (O’Donnell & Grace, 1995). Thus, stimulation of the fornix induced bistable cells to switch to the depolarized (active) state, and following transection of the fornix, no bistable cells were observed. More recently, stimulation of BLA afferents has been shown to result in a (comparatively brief) transition to the up state in accumbal neurons (Grace, 2000). Thus, it seems that the glutamatergic afferents to the NA are all sensitive to aspects of motivationally significant stimuli and send excitatory inputs to the NA that are integrated in the production of goal-directed behavior. Consistent with this notion are data demonstrating that glutamate release is responsible for the excitation of medium spiny neurons that occur in response to somatosensory stimuli and during behavior (Calabresi, Pisani, Centonze, & Bernardi, 1997; Wilson & Kawaguchi, 1996).
When considered as a whole, the electrophysiological and behavioral data suggest that activation of glutamatergic afferents to the accumbens produces a state of behavioral activation. Thus, activation of locomotor and goal-directed behavior seems to depend upon glutamatergic input to the NA. Within this context, it seems that the function of glutamate is to activate the production of motivated behavior. Remember that activating or energizing behavior was one of the principal features of motivation. Presumably, the direction is coded in the source of the glutamatergic input that is activated by the eliciting stimulus. Because the PFC, BLA, and hippocampus have been associated with different functional inputs (Kalivas et al., 1993a; Mogenson, Brudzynski, Wu, Yang, & Yim, 1993), it seems likely that these inputs code the memory and motor patterns activated by motivationally significant stimuli.
Glutamate and Plasticity
Although a primary function of glutamate within the motive circuit is activation of preexisting goal-directed behavior, glutamate has a clear and well-characterized role in synaptic and behavioral plasticity. Tetanic stimulation produces both long-term depression or potentiation in various nuclei in motive circuitry, and most studies reveal that these events are blocked by antagonizing excitatory amino acid transmission (Calabresi, Centonze, Gubellini, Marfia, & Benardi, 1999; Geiger et al., 1995). It is significant that recent studies have demonstrated modulation of excitatory transmission-based synaptic plasticity by mesocorticolimbic dopamine transmission (Bonci & Malenka, 1999; Thomas, Malenka, & Bonci, 2000), especially with regard to corticostriatal glutamatergic afferents (Calabresi, de Murtas, & Bernardi, 1997). Consistent with a role for glutamate in behavioral plasticity are data demonstrating that the initiation of long-term behavioral changes associated with chronic drug exposure for cocaine, amphetamine, and opioids is dependent upon glutamate receptor stimulation (Cornish & Kalivas, 2000; Wolf, 1998). Most consistently, glutamatergic neuroplasticity involves NMDA receptor stimulation, although an increasing role for mGluRs—perhaps in concert with iGluR activation—is being recognized (Anwyl, 1999).
GABA and Motivated Behavior
Behavioral studies, although they are not numerous, suggest an important role for GABA in the control of goal-directed behavior. Like glutamate, GABA is widely distributed throughout the brain and in vivo neurochemical evaluation of GABA transmission is difficult, making interpretations problematic. It is the most abundant inhibitory neurotransmitter and hyperpolarizes both projection and interneurons (Calabresi, Mercuri, de Murtas, & Bernardi, 1990; M. W. Jones, Kilpatrick, & Phillipson, 1988; Napier, Simson, & Givens, 1991). Consistent with the role of GABA as an inhibitory transmitter, stimulation of GABA receptors in the NA results in a decrease in firing frequency of neurons throughout the motive circuit. In some instances, biphasic effects are observed in projection cells, which results from relatively greater efficacy of GABA agonists or GABA receptor subtypes on GABAergic inhibitory interneurons (Grace & Bunney, 1985). Thus, low levels of GABAergic tone serve to stimulate projection neurons via inhibiting GABAergic interneurons, whereas greater levels of GABAergic tone directly hyperpolarize the projection cells.
Modulating GABA Transmission in the Motive Circuit
Consistent with the role of GABA as an inhibitory transmitter, electrical or pharmacological stimulation of the NA activates GABAergic projection cells, resulting in the suppression of firing in pallidal neurons that is reversed by application of GABA antagonists (D. L. Jones & Mogenson, 1980; Lamour, Dutar, Rascol, & Jobert, 1986). Similarly, stimulation of the VTAinhibits the firing of PFC neurons in a GABA- (and DA-) dependent fashion (Schilstrom et al., 2000). Stimulation of neurons in the NA that project to the VTA produces a more complicated effect on neurons in the VTA, where both excitation of dopamine cells by disinhibition and direct inhibition of secondary (GABAergic) cells is observed (Kalivas, 1993).
The most well-developed pharmacological literature regarding the effects of GABA on motivational circuitry is based on the role of GABAin regulating locomotor activity. In general, stimulating either GABAA or GABAB receptor subtypes produces a decrease in motor activity in most nuclei in the motive circuit. Thus, stimulating GABAB receptors in the VTA inhibits motor activity, whereas stimulation of GABAA receptors in the NA or VP inhibits activity (Kalivas, 1993). A notable exception to these observations is the effect of GABAB receptor activation in the MD, where a potent locomotor stimulation is observed (Churchill, Zahm, Duffy, & Kalivas, 1996). Thus, inhibition of the glutamatergic projection from the MD to the PFC promotes locomotor activity, an effect mediated in part by increasing mesoaccumbens dopamine transmission (Chuchill et al., 1996; M. W. Jones et al., 1988).
Direct infusion of either the GABAA agonist muscimol or the GABAB agonist baclofen into the NAS elicits a feeding behavior in satiated rats (Stratford & Kelley, 1997). These effects are probably due to presynaptic inhibition of excitatory glutamatergic drive because baclofen has been shown to inhibit glutamate release in the NA (Uchimura & North, 1991). It has also been demonstrated in striatal spiny neurons that baclofen administration inhibits stimulation-induced excitatory postsynaptic potentials without affecting resting membrane potential, again indicating a presynaptic GABABmediated effect on glutamate release (Nisenbaum, Berger, & Grace, 1993). These data suggest that either direct postsynaptic inhibition of NA neurons (GABAA) or presynaptic inhibition of glutamate release (GABAB) can elicit feeding behavior, indicative of a modulatory role for GABA in maintaining tone within the accumbens on the production of behavior. Consistent with this explanation, blockade of GABAA receptors in the VPm (which would be functionally equivalent to inhibition of GABA projection neurons within the NAs) has also been shown to elicit feeding behavior (Stratford, Kelley, & Simansky, 1999).
GABA receptor activation also appears to have a role in modulating drug self-administration. Thus, baclofen administration into the NA has been shown to inhibit heroin selfadministration (Xi & Stein, 1999), whereas administration of muscimol into the VTA blocks ethanol self-administration (Hodge, Hraguchi, Chappelle, & Samson, 1996). Similarly, the systemic administration of baclofen inhibits cocaine selfadministration (Brebner, Phelan, & Roberts, 2000; Roberts & Andrews, 1997).
Considering the pharmacological data previously outlined, it can be concluded that GABA has a modulatory role in the production of goal-directed behavior. Moreover, in general increasing GABA transmission appears to diminish motivated behavior, a notable exception being in the MD or GABA-mediated disinhibition of dopamine cell firing.
GABA and the Binding of Motivational Information
One necessary function of any circuit designed to integrate information and guide behavior is to provide a means to bind information over time. The binding of information in motivational circuitry permits an organism to persevere in goaldirected behavior in the presence of changes in external and internal stimuli. Similar binding of information has been characterized in visual circuitry in which topographically organized feedback loops are proposed to sustain information over time and facilitate the integration of previous visual stimuli with immediate stimuli, thereby permitting a coherent flow of information over time (Herrnstein, 1971; Yeomans, 1990). It has been proposed that such a topographically organized reciprocal feedback loop in motivational circuitry is formed by part of the limbic subcircuit consisting of the VTA, NAs, and VPm (see Figure 14.2, Panel A; Kalivas et al., 1993a). As described previously, tone within this subcircuit is maintained by reciprocal GABAergic interconections between these nuclei, and motivated behaviors can be disrupted or promoted by pharmacologically manipulating GABA transmission in any of these nuclei.
GABAergic Interconnection Between the Nucleus Accumbens and Ventral Pallidum
A difficulty arises in interpreting the effects of GABA on behavior when considering the effects of GABAergic tone in the NA and VP. Thus, one would expect inhibition of NA neurons to decrease activity in the GABAergic projection to the VP, thereby increasing activity within the VP via disinhibition (Mogenson et al., 1993). However, inhibition of the VP generally causes the same decrease in behavioral responding seen following inhibition of the NA. Thus, drug administration that results in opposite effects on neuronal activity in the VP produces very similar effects on responding.
This paradox is most apparent when examining the effects of GABA function on locomotor activity. Thus, treatments in the NAthat increase motor activity are generally known to be excitatory on spiny cells projecting to the VP, including glutamate receptor agonists, nicotinic agonists, GABA antagonists, and D1 agonists (Austin & Kalivas, 1988; Clarke & White, 1987; Willins, Narayanan, Wallace, & Uretsky, 1993). However, other treatments known to hyperpolarize spiny cells such as D2 and mu opioid agonists also elicit motor activation when microinjected into the NA (Clarke & White, 1987; Hakan & Eyl, 1995). It is noteworthy, however, that the increase in motor activity by D2 agonists in the NA is statedependent and is only observed in animals with an initial high level of activity (Brudzynski, Wu, & Mogenson, 1993).Also, the motor stimulant response elicited by mu opioid receptor activation is delayed and generally manifested 20–30 min after focal microinjection into the NA (Vezina, Kalivas, & Stewart, 1987). Thus, in contrast to the drugs that electrophysiologically activate spiny cell firing and produce rapid activation of motor activity, those drugs that acutely inhibit cell firing have a state-dependent or modulatory role that may not be indicative of the acute pharmacological inhibition.
Regardless of the ultimate effect the drugs have on the firing frequency of spiny cells, in every instance examined, stimulating GABAA receptors in the VPprevents the stimulation of motor activity by simultaneous drug administration into the NA. Moreover, the infusion of muscimol into the VP by itself has been shown to produce a decrease (Austin & Kalivas, 1990), no effect (Shreve & Uretsky, 1990), or an increase (Baud, Mayo, le Moal, & Simon, 1989) in locomotor activity—depending on the dose administered. These data seem to indicate that whether motor behavior is induced depends more on the fact that a drug changes ongoing GABAergic tone in the NA to VP projection, regardless of whether the change is an increase or decrease in tone.
Thus, the regulation of behavior by the GABAergic projection from the NAto VP can be excitatory or inhibitory, depending upon the situation. This speaks to a modulatory role for GABA in which either an increase or decrease in background tone can have effects on behavioral responsivity. The state-dependent effects of GABAtransmission may be in part due to the fact that spiny cells colocalize GABA with a number of different neuropeptides. Thus, divergent behavioral outcomes could potentially be due to differential activity of various peptides.
GABA-Neuropeptide Colocalization and Valence
There are a variety of neuropeptides within the motive circuit that colocalize with the more traditional transmitters already discussed. Because of their topographical distribution and interaction with these transmitter systems, investigators have examined a role for neuropeptides in the production of goaldirected behavior. The most well-described neuropeptide projections arise from the striatum (including the NA). About 90–95% of striatal neurons are GABAergic projection neurons. These projections neurons can be divided into two subtypes depending upon their projection targets, expression of DA receptors and peptide content. One subtype projects to the SN (striatonigral neurons), whereas the other projects to theVP (striatopallidal neurons; Gerfen, 1993; Kawaguchi, Wilson, & Emson, 1990). These have been termed the direct and indirect pathways, respectively (Albin, Young, & Penny, 1989).
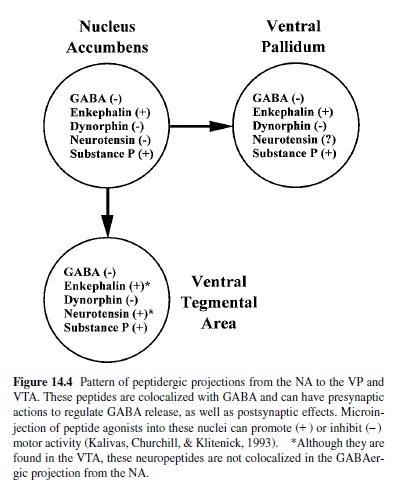
These two classes of neurons are differentially affected by dopamine based on the classes of DA receptor subtypes that they express. Striatonigral neurons mainly express D1 receptors that are positively coupled to adenylate cyclase, whereas striatopallidal neurons express D2 receptors that are negatively coupled to adenylate cyclase activity (Hersch et al., 1995; Le Moine & Bloch, 1995). Discretion in peptide localization is also observed in the striatonigral neurons that express dynorphin and substance P, whereas striatopallidal neurons express enkephalin and neurotensin (Beckstead & Kersey, 1985; Fallon & Leslie, 1986). It is notable that in the more ventral part of the striatum—especially in the shell of the NA—the projection to the VP is more promiscuous with respect to peptide content. Thus the NA to VP projection contains D1 receptors, substance P, and dynorphin, as well as enkephalin, neurotensin, and D2 receptors (Lu, Ghasemzadeh, & Kalivas, 1998). In contrast, the neurons projecting from the NAs to the VTA do not contain D2 receptors or enkephalin. Figure 14.4 illustrates this projection and also indicates whether stimulation of various peptidergic receptors activates (+ ) or inhibits (–) motor activity in the NA, VTA, or VP.
It seems plausible that the differential distribution of neuropeptides within the spiny cells of the NA reflects a functional difference in the regulation of goal-directed behavior. Consistent with this notion are data showing that the level of DAexpression affects the level of peptide expression. For example, 6-hydroxydopamine depletion leads to a decrease in substance P and dynorphin expression in nigrostriatal neurons but results in increased levels of enkephalin expression in striatopallidal neurons (Gerfen et al., 1990; Engber, Boldry, Kuo, & Chase, 1992). Additionally, transgenic animals lacking D1 receptors show decreased expression of substance P and dynorphin, with minimal or no effect on enkephalin expression (Drago, 1994; Drago, Gerfen, Westphal, & Steiner, 1996; Xu et al., 1994). On the other hand, mice that lack D2 receptors show mostly increased enpkephalin expression (Baik et al., 1995). Thus, neuropeptide levels in the striatum are dynamically and differentially regulated by the amount of DAreceptor activation.
Not only can DA function affect peptides, but peptide activity can also regulate the function of striatal projection neurons. Dynorphin receptor stimulation in the striatum inhibits the expression of immediate early genes (IEGs) typically induced by cocaine or D1 receptor agonists (Steiner & Gerfen, 1998). These results indicate that dynorphin acts as a negative feedback mechanism that regulates the activity of striatonigral neurons. In contrast, enkephalin receptor stimulation inhibits the expression of IEGs normally elicited by D2 receptor blockade, indicating that enkephalin acts as a negative feedback mechanism regulating striatopallidal neurons. We find it interesting that these adaptive changes in gene regulation are induced by situations that are also known to alter motivated behavior—chronic drug use (dynorphin) and chronic loss of D2 receptor function (enkephalin).
From the previous discussion, it seems that different classes of peptides can have opposing neuronal effects. This opposition seems to hold true for their electrophysiological effects as well. Iontophoretic application of morphine (a mu opioid receptor agonist) or enkephalin (a delta opioid receptor agonist) into the VTA increased the firing rate of mesolimbic DA neurons (Gysling & Wang, 1983). In contrast, kappa opioid receptor agonists decrease neuronal firing rates (Walker, Thompson, Frascella, & Friederich, 1987). These data suggest that mu and kappa opiates have opposing effects on dopaminergic activity, in addition to being differentially regulated be DA receptor activity.
Opioid peptides also have opposite roles in the regulation of certain types of behavioral learning that depend on affective valence. When a novel environment is repeatedly paired with a positive outcome, animals form a preference for that location and choose to spend time in it over an environment paired with a neutral outcome. Similarly, if an environment is paired with an aversive outcome, subjects avoid it when given a choice between that environment and a neutral one. Research has demonstrated that mu receptor agonists elicit place preferences, whereas kappa agonists elicit place aversions. Similarly, animals form a preference for tastes associated mu agonists and an aversion to tastes paired with kappa agonists (Mucha & Herz, 1985). These data indicate that mu activation produces a positive affective state, whereas kappa activation produces a negative one. This suggestion is consistent with reports from human subjects in which dynorphin (and other kappa agonists) has been shown to elicit dysphoria (Pfeiffer, Brandt, & Herz, 1986).
Neuropeptides have similarly been shown to regulate the production of goal-directed behaviors like cocaine seeking. Thus, with repeated exposure to cocaine, animals show an augmented responsivity to its behavioral activating effects. Enkephalin facilitates this phenomenon (Sala et al., 1995), whereas it is attenuated by kappa agonists (Heidbreder, Bobovic-Vuksanovic, Shoaib, & Shippenberg, 1995; Heidbreder, Goldberg, & Shippenberg, 1993). Furthermore, enkephalin and cocaine produce cross-sensitization when infused into the VTA (DuMars, Rodger, & Kalivas, 1988). In fact, repeated administration of enkephalin produced behavioral sensitization (Kalivas, Taylor, & Miller, 1985). Alternatively, the kappa agonist, U69593, blocks the reinstatement of coaine-seeking behavior following a period of drug abstinence (Schenk, Partridge, & Shippenberg, 2000).
Infusion of substance P, the enkephalin analog D-Ala(2)methionine(2)-enkephalinamide, or mu or delta opioid receptor agonists into the VTA produces locomotor activity (Joyce & Iversen, 1979; Joyce, Koob, Strecker, Iversen, & Bloom, 1981; Kalivas, Widerlov, Stanley, Breese, & Prange, Jr., 1983). Additionally, enkephalin infusion into the NA or SN facilitates spontaneous motor activity (Kalivas, 1985). However kappa opioid agonists attenuate the locomotor activation produced by morphine (Pearl & Glick, 1996), once again suggesting an opposite modulation of behavioral output by neuropeptides.
When considered as a whole, neuropeptides within the motive circuit clearly play a role in regulating behavioral output. Furthermore, whereas mu or delta opioid receptor activation seems to play a generally facilitory role, kappa opioid receptor activation seems to be primarily inhibitory. Thus, at least with the opioid peptides it is plausible that an important function of these peptides within the motive circuit is to provide valence. However, the neurobiological underpinnings to evaluate this hypothesis are largely missing. For example, it is not clear under what electrophysiological or behavioral conditions are the cotransmitted peptides released with or in lieu of GABA. Moreover, in certain instances, notably mu opioid receptor stimulation in the VP, the neuropeptide has both a pre- and postsynaptic action producing opposite effects on projection cells (Napier & Mitrovic, 1999; Olive, Anton, Micevych, Evans, & Maidment, 1997), and it is unclear under what circumstances one or the other receptor population is preferentially activated.
Summary
This review has endeavored to outline the basic circuitry most consistently identified as being important in regulating motivation. It is notable that the circuit contains two overlapping orientations: (a) cortical and subcortical circuitry and (b) limbic and motor circuitry. Moreover, at the outset we hypothesized distinct roles for the major neurotransmitters within the circuit, including dopamine, glutamate, GABA, and various neuropeptides. In this summary we briefly revisit the literature according to these organizing principles.
Cortical and Subcortical Circuitry
Figure 14.2 shows that there exists an interconnected circuit between subcortical nuclei such as the VTA, NA, VP, and MD, and cortical or allocortical structures such as the PFC, amygdala, and hippocampus. To some extent the relationship between the cortical and subcortical nuclei is rectified in that the flow of information is more direct from the cortex to subcortical nuclei than from the subcortical nuclei back to the cortex (the notable exception being the VTA, which projects to all cortical regions). Based upon this anatomy and the contributions that all three cortical regions provide in terms of cognition and memory, it can be proposed that the cortical regions imbue the motivational state with memories linked to the environmental stimulus, ranging from conditioned behavioral responses to salient stimuli provided by the amygdala to incorporating recent information stored as working memory by the prefrontal cortex. The information transmitted by these glutamatergic cortical afferents is integrated in the subcortical nuclei and directed into motor circuitry to facilitate adaptive behavioral responding. Of course, memories are most relevant in guiding behavioral responding to familiar stimuli— and the more novel a stimulus, the less valuable previous experiences will be in generating appropriate behavioral responses. In situations of high novelty, behavior appears to be guided more by subcortical regions of motivational circuitry. Thus, exploratory behavior in response to a novel environment is disrupted most effectively by pharmacological interventions in the VTA, NA, or VP (Hooks & Kalivas, 1995). It is notable that numerous data support a role for dopamine neurons in the VTA in organizing or signaling behavioral responses to novel stimuli (Schultz, 1998).
Motor Versus Limbic Circuitry
The fact that there are two separate, but interactive subcircuits within the motive circuit suggests that they have separable functions in the production of goal-directed behaviors. Limbic structures, like the VTA, BLA, and hippocampus are more intimately connected with the PFCv, NAs, and VPm, whereas motor structures like primary motor cortex, SN, and the PPN are more intimately connected with the PFCd, NAc, and VPl. This leads to the suggestion that the motor loop is more directly involved in sending information about the well-learned responses to motor systems (i.e., procedural memory), whereas the limbic loop is more directly involved in learning about motivationally relevant stimuli and subsequently integrating incoming information about such stimuli when they are presented. This role for the limbic loop is consistent with information showing that animals will learn to self-administer a variety of drugs directly into the NAs but not the NAc (for a review, see McBride, Murphy, & Ikemoto, 1999), suggesting that the learning necessary for animals to form an association between their behavior and intracranial drug delivery requires the involvement of limbic processing.
A potential role for the motor loop in the performance of procedural memory is consistent with recent evidence from our laboratory showing that cocaine-induced reinstatement of drug-seeking behavior is mediated by a series circuit from the PFCd through the NAc and then out to the motor system via the VPl (McFarland & Kalivas, 2001). Thus, well-learned behaviors are relatively automatic and can be elicited independent of limbic input. However, reinstatement elicited by drug-predictive cues has been shown to critically depend upon the BLA, a limbic structure (Meil & See, 1997; Grimm & See, 2000), indicating that multiple structures can be involved in the production of behavior. The limbic structure(s) most critical would depend upon the type of sensory and memory processing required for production of the goaldirected behavior. Thus, a stimulus with motivational significance triggers behavior within the limbic subcircuit to the motor subcircuit to initiate the behavioral output.
Within this framework, the PFCd plays a particularly important role in the initiation of well-learned responses. The PFCd is a primary projection target of the MD, which forms a rectified bridge between the limbic and motor loops, presumably allowing processing to move from limbic to motor subcircuits. Consistent with an important role by the PFCd are data demonstrating that damage to the PFC results in deficits in response inhibition (e.g., Bussey, Muir, Everitt, & Robbins, 1997; Roberts & Wallis, 2000; Seamans, Floresco, & Phillips, 1998). Electrophysiological studies examining neuronal activation patterns within the PFC have found that some neurons consistently exhibit altered firing rates immediately preceding the emission of lever-press behavior resulting in delivery of cocaine or heroin (Chang, Zhang, Janak, & Woodward, 1997; Chang, Janak, & Woodward, 2000). It is interesting that many of these neurons show correlated activity with NA neurons. These data are consistent with the notion that the PFC plays an important role in response initiation. Furthermore, in human subjects, regional activation studies have shown that the prefrontal cortex is associated with preparation of motor responses and inhibition of inappropriate responses (Chang et al., 2000; de Zubicaray, Zelaya, Andrew, Williams, & Bullmore, 2000).
Functions for Neurotransmitters in the Motive Circuit
Various hypotheses were proposed at the outset of this review regarding the role of each transmitter. The following summary evaluates the veracity of each hypotheses based upon the key findings previously described.
- Glutamate stimulates behavior, and the anatomical origin of the activated glutamatergic afferents provides motor memory to provoke the appropriate behavioral response. In addition, under the appropriate conditions glutamate transmission promotes neuroplasticity permitting learning and behavioral adaptations to occur. This hypothesis is directly supported by the anatomical organization of glutamate projections in the motive circuit in that all glutamate derives from cortical and allocortical structures or within thalamic projections to the cortex. Thus, glutamate is involved primarily in driving and maintaining learned behavioral responses to familiar motivationally relevant stimuli. Indeed, at a cellular level, learned behaviors are thought to great extent to be encoded in changes in excitatory transmission, indicating that neuroadaptations in glutamate transmission contribute to cellular memory. However, in addition to mediating established behaviors, substantial data indicate that glutamate is critical for initial neuroplastic events associated with establishing adaptive behavioral responses. Thus, many changes in gene expression elicited by novel motivational stimuli depend on glutamate transmission, and glutamate receptor antagonists prevent the enduring establishment of adaptive behavioral responses.
- Dopamine supports plasticity and learning by engaging the appropriate cellular machinery to modify neuronal communication, especially excitatory trasmission. Dopamine is the most widely studied neurotransmitter in regulating motivation and at various times has been assigned a primary role in most aspects of motivation. Considering the data outlined previously, it can be argued that the primary role for dopamine is in the acquisition of adaptive behavioral responses to motivationally relevant behaviors, with a lesser role in the maintenance of behavioral responding. Thus, dopamine neurons are activated in response to novel, motivationally relevant stimuli and by stimuli that signal an impending reward. In contrast, dopamine has little role in carrying out a well-established adaptive behavioral response.
- GABA regulates overall circuit tone and thereby serves to bindorsustainananimal’smotivationalstateuntilthegoal object can be achieved.The most striking anatomical characteristic of GABA neurons within the circuit is the triad of subcortical nuclei that are interconnected by GABAergic projections, including the VTA, NA, and VP.As described previously, this subcortical triad is not involved in the longterm storage of information required for eliciting established adaptive responses. However, it is critical for the execution of both new and established behavioral responses to motivationally relevant stimuli. GABA transmission within each nucleus is responsible for maintaining these behaviors over Given the topographic connectivity of the GABA projections, this triad has the potential to provide background tone or state-dependence of responding to motivationally relevant stimuli. Thus, under conditions of high GABA tone, less vigorous behavioral responses would be expected and vice versa. A gradual shifting in GABAergic tone depending on the changing temporal quality of a stimulus would act as an information buffer to maintain continuity of the motivational state over time.
- Neuropeptides contribute to subjective valence. The study of neuropeptides has been the least informative to a unified hypothesis of how they may function in the motive circuit. The colocalization of neuropeptides in GABAergic neurons in the NA has been the most carefully studied. At least with regards to the behavioral effects of enkephalin and dynorphin in the NA, the data support a role for the neuropeptides in providing valence to a given stimulus. However, the overall dearth of knowledge regarding the cellular and behavioral regulation of peptide release makes an overarching hypothesis of neuropeptide function impossible to evaluate. Indeed, given the wide variety of neuropeptides within the motive circuit, it seems unlikely that any overarching hypothesis encompassing all of the peptides will be validated.
Conclusions
Although we have now achieved a state of knowledge permitting some general characteristics to be assigned to motivational circuitry, emerging subtleties in how the circuit functions will profoundly impact the generalizations outlined previously. Significant sources of information currently emerging are in at least two arenas:
- In vivo electrophysiological studies that permit the evaluation of ensembles of neurons within a single nucleus or between multiple nuclei will provide direct testing of the postulated roles assigned to the nuclei and transmitters within the circuit. However, the data to date reveal substantial heterogeneity in neuronal responses to various motivationally relevant stimuli, indicating that a large amount of data will be required to accurately describe the patterns of interactions within the circuit.
- Establishing behavioral responses to motivationally relevant stimuli requires both short-term and long-term neuroplasticity. The neurosciences are currently engaged in a revolution affecting our understanding of how changes in gene expression and protein trafficking mediate synaptic plasticity. As this process becomes better understood, it will be applied to behavioral plasticity and the neuroscienceswilllikelyexperiencealargeleapforwardincomprehending the role of motive circuitry in the acquisition of behavioral responding for motivationally relevant stimuli.
Bibliography:
- Aggleton, J. P., Burton, M. J., & Passingham, R. E. (1980). Cortical and subcortical afferents to the amygdala in the rhesus monkey (Macaca mulatta). Brain Research, 190, 347–368.
- Albin, R. L., Young, A. B., & Penny, J. B. (1989). The functional anatomy of basal ganglia disorders. Trends in Neuroscience, 12, 366–375.
- Anden, N. E., Fuxe, K., Hamberger, B., & Hokfelt, T. (1996). A quantitative study on the nigro-neostriatal dopamine neurons. Acta Physiologica Scandinavica, 67, 306–312.
- Anwyl, R. (1999). Metabotropic glutamate receptors: Electrophysiological properties and role in plasticity. Brain Research Reviews, 29, 83–120.
- Attarian,S.,&Amalric,M.(1997).Microinfusionofthemetabotropic glutamate receptor agonist 1S,3R-1-aminocyclopentane-1,3dicarboxylic acid into the nucleus accumbens induces dopaminedependent locomotor activation in the rat. European Journal of Neuroscience, 9, 809–816.
- Austin, M. C., & Kalivas, P. W. (1988). The effect of cholinergic stimulation in the nucleus accumbens on locomotor behaivor. Brain Research, 441, 209–214.
- Austin, M. C., & Kalivas, P. W. (1990). Enkephalinergic and GABAergic modulation of motor activity in the ventral pallidum. Journal of Pharmacology and Experimental Therapeutics, 252, 1370–1377.
- Bacon, S. J., Headlam, A. J., Gabbott, P. L., & Smith, A. D. (1996). Amygdala input to medial prefrontal cortex (mPFC) in the rat: A light and electron microscope study. Brain Research, 720, 211–219.
- Baik, J.-H., Picetti, R., Saiardi, A., Thiriet, G., Dierich, A., Depaulis, A., et al. (1995). Parkonsonian-like motor impairment in mice lacking dopamine D2 Nature, 377, 424–428.
- Baud, P., Mayo, W., le Moal, M., & Simon, H. (1989). Locomotor hyperactivity in the rat after infusion of muscimol and [D-Ala2]Met-enkephalin into the nucleus basalis magnocellularis: Possible interaction with cortical cholinergic projections. Brain Research, 452, 203–211.
- Bazhenov, M., Timofeev, I., Steriade, M., & Sejnowski, T. J. (1998). Cellular and network models for intrathalamic augmenting responses during 10-Hz stimulation. Journal of Neurophysiology, 79, 2730–2748.
- Beckstead, R. M., Domesick, V. B., & Nauta, W. J. H. (1979). Efferent connections of the substantia nigra and ventral tegmental area in the rat. Brain Research, 175, 191–217.
- Beckstead, R. M., & Kersey, K. S. (1985). Immunohistochemical demonstration of differential substance P-, Met-enkephalin, and glutamic acid decarboxylase-containing cell and axon distributions in the corpus striatum of the cat. The Journal of Comparative Neurology, 232, 481–498.
- Beninger, R. J. (1983). The role of dopamine in locomtor activity and learning. Brain Research Reviews, 6, 173–196.
- Beninger,R.J.,&Hahn,B.L.(1983).Pimozideblocksestablishment but not expression of amphetamine-produced environmentspecific conditioning. Science, 220, 1300–1304.
- Beninger, R. J., & Herz, R. S. (1986). Pimozide blocks establishment but not expression of cocaine-produced environmentspecific conditioning. Life Sciences, 38, 1425–1430.
- Berendse, H. K., Galis-de Graaf, Y., & Groenewegen, H. J. (1992). Topographical organization and relationship with ventral striatal compartments of prefrontal corticostriatal projections on the rat. The Journal of Comparative Neurology, 316, 314– 347.
- Bindra, D. (Ed.). (1969). The interrelated mechanisms of reinforcement and motivation, and the nature of their influence on response. Lincoln: University of Nebraska Press.
- Bliss, T. V. P., & Collingridge, G. L. (1993). A synaptic model of memory: Long-term potentiation in the hippocampus. Nature, 361, 31–39.
- Boldry, R., Willins, D., Wallace, L., & Uretsky, N. (1991). The role of endogenous dopamine in the hypermotility response to intraaccumbens AMPA. Brain Research, 559, 100–108.
- Bonci, A., & Malenka, R. C. (1999). Properties and plasticity of excitatory synapses on dopaminergic and GABAergic cells in the ventral tegmental area. Neuroscience, 19, 3723–3730.
- Bowers, W., Hamilton, M., Zacharo, R. M., & Anisman, H. (1985). Differential effects of pimozide on response-rate and choice accuracy in a self-stimulation paradigm in mice. Pharmacology, Biochemistry, and Behavior, 22, 512–526.
- Bozarth, M. A., & Wise, R. A. (1981). Heroin reward is dependent on a dopaminergic substrate. Life Sciences, 29, 1881–1886.
- Brebner, K., Phelan, R., & Roberts, D. (2000). Effect of baclofen on cocaine self-administration in rats reinforced under fixedratio 1 and progressive-ratio schedules. Psychopharmacology, 148(3), 314–321.
- Brinley-Reed, M., & McDonald, A. J. (1999). Evidence that dopaminergic axons provide a dense innervation of specific neuronal subpopulations in the rat basolateral amygdala. Brain Research, 850, 127–135.
- Brudzynski, S. M., Wu, M., & Mogenson, G. J. (1993). Decreases in rat locomotor activity as a result of changes in synaptic transmission to neurons within the mesencephalic locomotor region. Canadian Journal of Physiological Pharmacology, 71, 394–406.
- Bussey, T. J., Muir, J. L., Everitt, B. J., & Robbins, T. W. (1997). Triple dissociation of anterior cingulate, posterior cingulate, and medial frontal cortices on visual discrimination tasks using a touchscreen testing procedure for the rat. Behavioral Neuroscience, 111, 920–936.
- Calabresi, P., Centonze, D., Gubellini, P., Marfia, G., & Bernardi, G. (1999). Glutamate-triggered events inducing corticostriatal long-term depression. The Journal of Neuroscience, 19, 6102– 6110.
- Calabresi, P., De Murtas, M., & Bernardi, G. (1997). The neostriatum beyond the motor function: Experimental and clinical evidence. Neuroscience, 78, 39–60.
- Calabresi, P., Mercuri, N. B., de Murtas, M., & Bernardi, G. (1990). Endogenous GABA mediates presynaptic inhibition of spontaneous and evoked excitatory synaptic potentials in the rat neostriatum. Neuroscience Letters, 118, 99–102.
- Calabresi, P., Pisani, A., Centonze, D., & Bernardi, G. (1997). Synaptic plasticity and physiological interactions between dopamine and glutamate in the striatum. Neuroscience and Behavioral Reviews, 21, 519–523.
- Carr, D. B., O’Donnell, P., Card, J., & Sesack, S. (1999). Dopamine terminals in the rat prefrontal cortex synapse on pyramidal cells that project to the nucleus accumbens. The Journal of Neuroscience, 19, 11049–11060.
- Carr, D. B., & Sesack, S. R. (1996). Hippocampal afferents to the rat prefrontal cortex: Synaptic targets and relation to dopamine terminals. The Journal of Comparative Neurology, 369(1), 1–15.
- Carr, D. B., & Sesack, S. R. (2000a). GABA-containing neurons in the rat ventral tegmental area project to the prefrontal cortex. Synapse, 38(2), 114–123.
- Carr, D. B., & Sesack, S. R. (2000b). Projections from the rat prefrontal cortex to the ventral tegmental area: Target specificity in the synaptic associations with mesoaccumbens and mesocortical neurons. The Journal of Neuroscience, 20, 3864–3873.
- Chang, J. Y., Janak, P. H., & Woodward, D. J. (2000). Neuronal and behavioral correlations in the medial prefrontal cortex and nucleus accumbens during cocaine self-administration by rats. Neuroscience, 99, 433–443.
- Chang, J. Y., Zhang, L., Janak, P. H., & Woodward, D. J. (1997). Neuronal responses in prefrontal cortex and nucleus accumbens during heroin self-administration in freely moving rats. Brain Research, 754, 12–20.
- Christie, M. J., Summers, R. J., Stephenson, J. A., Cook, C. J., & Beart, P. M. (1987). Excitatory amino acid projections to the nucleus accumbens septi in the rat: Aretrograde transport study utilizing D[3H]aspartate and [3H]GABA. Neuroscience, 22, 425–439.
- Churchill, L., Dilts, R., & Kalivas, P. (1990). Changes in y-aminobutyric acid,u-opioid and neurotensin receptors in the accumbens-pallidal projection after discrete quinolinic acid lesions in the nucleus accumbens. Brain Research, 511, 41–54.
- Churchill, L., & Kalivas, P. W. (1994). A topographically organized GABAergic projection from the ventral pallidum to the nucleus accumbens in the rat. The Journal of Comparative Neurology, 345, 579–595.
- Churchill, L., & Kalivas, P. W. (1999). The involvement of the mediodorsal nucleus of the thalamus and the midbrain extrapyramidal area in locomotion elicited from the ventral pallidum. Behavioural Brain Research, 104, 63–71.
- Churchill, L., Zahm, D. S., Duffy, T., & Kalivas, P. W. (1996). The mediodorsal nucleus of the thalamus: II. Behavioral and neurochemical effects of GABA agonists. Neuroscience, 70, 103–112.
- Clarke, D., & White, F. (1987). Review: D1 dopamine receptor—the search for a function: A critical evaluation of the D1/D2 dopamine receptor classification and its functional implications. Synapse, 1, 347–360.
- Corbett, D., & Wise, R. A. (1980). Intracranial self-stimulation in relation to the ascending dopminergic systems of the midbrain: A moveable microelectrode study. Brain Research, 185, 1–15.
- Cornish, J. L., Duffy, P., & Kalivas, P. W. (1999). A role of nucleus accumbens glutamate transmission in the relapse to cocaineseeking behavior. Neuroscience, 93, 1359–1368.
- Cornish, J. L., & Kalivas, P. W. (2000). Glutamate transmission in the nucleus accumbens mediates relapse in cocaine addiction. The Journal of Neuroscience, 20, 1–5.
- Davis, C. M., Moskovitz, B., Nguyen, M. A., Tran, B. B., Arai, A., Lynch, G., et al. (1997). A profile of the behavioral changes produced by facilitation of AMPA-type glutamate receptors. Psychopharmacology, 133, 161–167.
- de Wit, H., & Wise, R. A. (1977). Blockade of cocaine reinforcement in rats with the dopamine receptor blocker pimozide, but not with the noradrenergic blockers phenolamine or phenoxybenzamine. Canadian Journal of Psychology, 31, 195–203.
- de Zubicaray, G. I., Zelaya, F. O., Andrew, C., Williams, S. C., & Bullmore, E. T. (2000). Cerebral regions associated with verbal response initiation, suppression and strategy use. Neuropsychologia, 38, 1292–1304.
- di Chiara, G., Acquas, E., Tanda, G., & Cadoni, E. (1993). Drugs of abuse: Biochemical surrogates of specific aspects of natural reward? Biochemical Society Symposium, 59,
- Doherty, M. D., & Gratton, A. (1992). High speed chronoamperometric measurements of mesolimbic and nigrostriatal dopamine relelase associated with repeated daily stress. Brain Research, 586, 295–302.
- Donzanti, B. A., & Uretsky, N. J. (1983). Effects of excitatory amino acids in locomotor activity after bilateral microinjection into the rat nucleus accumbens: Possible dependence on dopaminergic mechanisms. Neuropharmacology, 22, 971–970.
- Drago, J. (1994). Altered striatal function in a mutant mouse lacking D1A dopamine receptors. Neurobiology, 91, 12564–12568.
- Drago, J., Gerfen, C. R., Westphal, H., & Steiner, H. (1996). D1 dopamine deficient mouse: Cocaine induced regulation of immediate early gene and substance Pexpression in the striatum. Neuroscience, 74, 813–823.
- DuMars, L. A., Rodger, L. D., & Kalivas, P. W. (1988). Behavioral cross-sensitization between cocaine and enkephalin in theA10 dopamine region. Behavioural Brain Research, 27, 87–91.
- Engber, T. M., Boldry, R. C., Kuo, S., & Chase, T. N. (1992). Dopaminergic modulation of striatal neuropeptides: Differential effects of D1 and D2 receptor stimulation on somatostatin, neuropeptide Y, neurotensin, dynorphin, enkephalin. Brain Research, 581, 261–268.
- Ettenberg, A. (1989). Dopamine, neuroleptics and reinforced behavior. Neuroscience and Behavioral Reviews, 13, 105–111.
- Ettenberg, A., & Horvitz, J. C. (1990). Pimozide prevents the response reinstating effects of water reinforcement in rats. Pharmacology, Biochemistry, and Behavior, 31, 861–865.
- Ettenberg,A., Pettit, H. O., Bloom, F. E., & Koob, G. F. (1982). Heroin and cocaine intravenous self-administration in rats: Mediation by separateneuralsystems.Psychopharmacology,78,204–209.
- Evenden, J. L., & Robbins, T. W. (1983). Dissociable effects of d-amphetamine, chlordiazepoxide, and alpha-fluphenthixol on choice and rate measures of reinforcement in the rat. Psychopharmacology, 79, 180–186.
- Everitt, B. J., & Robbins, T. W. (1992). Amygdala-ventral striatal interactions and reward-related processes. In J. P. Aggleton (Ed.), The amygdala: Neurobiological aspects of emotion, memory and mental dysfunction (pp. 401–429). New York: Wiley-Liss.
- Fallon, J. H., & Leslie, F. M. (1986). Distribution of dynorphin and enkephalin peptides in the rat brain. The Journal of Comparative Neurology, 249, 293–336.
- Fallon, J. H., & Moore, R. Y. (1978). Catecholamine innervation of basal forebrain: IV. Topography of the dopamine projection to the basal forebrain and striatum. The Journal of Comparative Neurology, 180, 545– 580.
- Fibiger, H. C., & Phillips, A. G. (1986). Reward, motivation, cognition: Psychobiology of mesotelencephalic dopamine systems. Handbook of Physiology I, 4, 647–675.
- Firth, C. D., Friston, K. J., Herold, S., Silbersweg, D., Fletcher, P., Cahill, C., et al. (1995). Regional brain activity in chronic schizophrenic patients during the preformance of a verbal fluency task. British Journal of Psychiatry, 167, 343–349.
- Fonnum, F., Myhrer, T., Paulsen, R. E., Wangen, K., & Oksengard, A. R. (1995). Role of glutamate receptors in memory function and Alzheimer’s disease. Annals of the New York Academy of Sciences, 757, 475–486.
- Fonnum, F., Storm-Mathiasen, J., & Divac, I. (1981). Bichemical evidence for glutamate as neurotransmitter in the cortico-striatal and cortico-thalamic fibres in rat brain. Neuroscience, 6, 863– 875.
- Fouriezos, G., Hansson, P., & Wise, R. A. (1978). Neurolepticinduced attenuation of brain stimulation reward. Journal of Comparative and Physiological Psychology, 92, 659–669.
- Franklin, K. B. J., & McCoy, S. N. (1979). Pimozide-induced extinction in rats: Stimulus control of responding rules out motor deficit. Pharmacology, Biochemistry, and Behavior, 11, 71–76.
- Füxe, K., Hökfelt, T., Johansson, O., Lidbrink, P., & Ljungdahl, A. (1974). The origin of the dopamine nerve terminals in limbic and prontal cortex: Evidence for meso-cortico dopamine neurons. Brain Research, 82, 349–355.
- Gallistel, C. R., Boytim, M., Gomita, Y., & Klebanoff, L. (1982). Does pimozide block the reinforcing effect of brain stimulation? Pharmacology, Biochemistry, and Behavior, 17, 769–781.
- Gasbarri, A., Verney, C., Innocenzi, R., Campana, E., & Pacitti, C. (1994). Mesolimbic dopaminergic neurons innervating the hippocampal formation in the rat: A combined retrograde tracing and immunohistochemical study. Brain Research, 668, 71–79.
- Geiger, J. R. P., Melcher, T., Koh, D.-S., Satmann, B., Seeburg, P. H., Jonas, P., et al. (1995). Relative abundance of subunit mRNAs determines gating and Ca2 permeability of AMPA receptors in principal neurons and interneurons in rat CNS. Neuron, 15, 193–204.
- Gerber, G. J., Sing, J., & Wise, R. A. (1981). Pimozide attenuates lever pressing for water in rats. Pharmacology, Biochemistry, and Behavior, 14, 201–205.
- Gerfen, C. R. (1993). The neostriatal mosaic: Multiple levels of compartmental organization in the basal ganglia. Annual Review of Neuroscience, 15, 285–320.
- Gerfen, C. R., Engbar, T. M., Mahan, L. C., Susel, Z., Chase, T. N., Monsma, F. J., Jr., et al. (1990). D1 and D2 dopamine receptorregulated gene expression of striatonigral and striatopallidal neurons. Science, 250, 1429–1432.
- Grace, A. (2000). Gating of information flow within the limbic system and the pathophysiology of schizophrenia. Brain Research Reviews, 31, 330–341.
- Grace, A. A., & Bunney, B. S. (1985). Opposing effects of striatonigral feedback pathways on midbrain dopamine cell activity. Brain Research, 333, 271–284.
- Gray, C. M., Maldonado, P. E., Wilson, M., & McNaughton, B. (1995). Tetrodes markedly improve the reliability and yield of single-unit isolation and multi-unit recording in cat striate cortex. Journal of Neuroscience Methods, 63, 43–54.
- Grimm, J., & See, R. (2000). Dissociation of primary and secondary reward-relevant limbic nuclei in an animal model of relapse. Neuropsychopharmacolgy, 22, 473–479.
- Groenewegen, H. J. (1988). Organization of afferent connections of the mediodorsal thalamic nucleus in the rat, related to mediodorsalprefrontal topography. Neuroscience, 24, 379–431.
- Groenewegen, H. J., Berendse, H. W., & Haber, S. N. (1993). Organization of the output of the ventral striatopallidal system in the rat: Ventral pallidal efferents. Neuroscience, 57, 113– 142.
- Groenewegen, H. J., Berendse, H. W., Wolters, J. G., & Lohman, A. H. (1990). The anatomical relationship of the prefrontal cortex with the striatopallidal system, the thalamus and the amygdala: Evidence for a parallel organization. Progress in Brain Research, 85, 95–116.
- Groenewegen, H. J., Vermeulen-Van der Zee, A., Te Kortschot, A., & Witter, M. P. (1987). Organization of the projections from the subiculum to the ventral striatum in the rat: A study using anterograde transport of Phaseolus vulgaris-leucoagglutinin. Neuroscience, 23, 103–112.
- Grove, E. A. (1988). Neural associations of the substantia innominata in the rat: Afferent connections. The Journal of Comparative Neurology, 277, 315–346.
- Gysling, K., & Wang, R. Y. (1983). Morphine-induced activation of A10 dopamine neurons in the rat brain. Brain Research, 277, 119–127.
- Haber, S., & Fudge, J. (1997). The primate substantia nigra and VTA: Integrative circuitry and function. Critical Reviews in Neurobiology, 11, 323–342.
- Haber, S., & Fudge, J. (2000). The central nucleus of the amygdala projection to dopamine subpopulations in primates. Neuroscience, 97, 479–494.
- Haber, S. N., Groenewegen, H. J., Grove, E. A., & Nauta, W. J. H. (1985). Efferent connections of the ventral pallidum: Evidence of a dual striatopallidofugal pathway. The Journal of Comparative Neurology, 235, 322–335.
- Hakan, R. L., & Eyl, C. (1995). Neuropharmacology of the nucleus accumbens: Iontophoretic applications of morphine and nicotine have contrasting effects on single-unit responses evoked by ventral pallidal and fimbria stimulations. Synapse, 20, 175–184.
- Hatfield, T., Han, J., M. C., Gallagher, M., & Holland, P. (1996). Neurotoxic lesions of basolateral, but not central amygdala interfere with Pavlovian second-order conditioning and reinforcer devaluation effects. The Journal of Neuroscience, 16, 5256–5265.
- Hauber, W., Bohn, I., & Giertler, C. (2000). NMDA, but not dopamine D2, receptors in the rat nucleus accumbens are involved in guidance of instrumental behavior by stimuli predicitng reward magnitude. The Journal of Neuroscience, 20, 6282–6288.
- Heidbreder, C. A., Bobovic-Vuksanovic, D., Shoaib, M., & Shippenberg, T. S. (1995). Development of behavioral sensitization to cocaine: Influence of kappa opioid receptor agonists. Journal of Pharmacology and Experimental Therapeutics, 275, 150–163.
- Heidbreder, C. A., Goldberg, S. R., & Shippenberg, T. S. (1993). The kappa-opioid receptor agonist U-69593 attenuates cocaineinduced behavioral sensitization in the rat. Brain Research, 616, 335–338.
- Heimer, L., Alheid, G. F., & Zahm, D. S. (1993). Basal forebrain organization: An anatomical framework for motor aspects of drive and motivation. In P. W. Kalivas & C. D. Barnes (Eds.), Limbic motor circuits and neuropsychiatry (pp. 1–32). Boca Raton, FL: CRC Press.
- Heimer, L., Zahm, D. S., Churchill, L., Kalivas, P.W., &Wohltmann, C. (1991). Specificity in the projection patterns of accumbal core and shell in the rat. Neuroscience, 41, 89–125.
- Hernandez-Lopez, S., Bargas, J., Surmeier, D. J., Reyes, A., & Galarraga, E. (1997). D1 receptor activation enhances evoked discharge in neostriatal medium spiny neurons by modulating an L-type Ca2 The Journal of Neuroscience, 17, 3334–3342.
- Herrnstein, R. J. (1971). Qunatitative hedonism. Journal of Psychiatric Research, 8,
- Hersch, S. M., Ciliax, B. J., Gutekunst, C.-A., Rees, H. D., Heilman, C. J., Yung, K. K. L., et al. (1995). Electron microscopic analysis of D1 and D2 dopamine receptor proteins in the dorsal striatum and their synaptic relationships with motor corticostriatal afferents. The Journal of Neuroscience, 15, 5222–5237.
- Herzog, A. G., & Van Hoesen, G. W. (1976). Temporal neocortical afferent connections to the amygdala in the rhesus monkey. Brain Research, 115, 57–69.
- Hitchcott, P., & Phillips, G. (1998). Double dissociation of the behavioral effects of R() 7-OH-DPAT infusions in the central and basolateral amygdala nuclei upon Pavlovian and instrumental conditioned appetitive behaviors. Psychopharmacology, 140, 458–469.
- Hodge, C., Hraguchi, M., Chappelle, A., & Samson, H. (1996). Effects of ventral tegmental micrinjections of the GABA-A agonist muscimol on self-administration of ethanol and sucrose. Pharmacology, Biochemistry, and Behavior, 53, 971–977.
- Hooks, M. S., & Kalivas, P. W. (1995). The role of mesoaccumbenspallidal circuitry in novelty-induced behavioral activation. Neuroscience, 64, 587–597.
- Horvitz, J. C., & Ettenberg, A. (1991). Conditioned incentive properties of a food-paired conditioned stimulus remain intact during dopamine receptor blockade. Behavioral Neuroscience, 105, 536–541.
- Jones, B., & Mishkin, M. (1972). Limbic lesions and the problem of stimulus-reinforcement associations. Experimental Neurology, 36, 362–377.
- Jones, D. L., & Mogenson, G. J. (1980). Nucleus accumbens to globus pallidus GABA projection: Electrophysiological and ionotophoretic investigations. Brain Research, 188, 93–105.
- Jones, M.W., Kilpatrick, I. C., & Phillipson, O.T. (1988). Dopamine function in the prefrontal cortex of the rat is sensitive to reduction of tonic GABA-mediated inhibition in the thalamic mediodorsal nucleus. Experimental Brain Research, 69, 623–634.
- Joyce, E. M., & Iversen, S. D. (1979). The effect of morphine applied locally to mesencephalic dopamine cell bodies on spontaneous motor activity in the rat. Neuroscience Letters, 14, 207– 212.
- Joyce, E. M., Koob, G. F., Strecker, R., Iversen, S. D., & Bloom, F. (1981). The behavioral effects of enkephalin analogues injected into the ventral tegmental area and globus pallidus. Brain Research, 221, 359–370.
- Kaatz, W., & Albin, R. L. (1995). Intrastriatal and intrasubthalamic stimulation of metabotropic glutamate receptors: Abehavioral and FOS immunohistochemical study. Neuroscience, 66, 55–65.
- Kalivas, P. W. (1985). Interactions between neuropeptides and dopamine neurons in the ventromedial mesencephalon. Neuroscience and Behavioral Reviews, 9, 573–587.
- Kalivas, P. W. (1993). Neurotransmitter regulation of dopamine neurons in the ventral tegmental area. Brain Research Reviews, 18, 75–113.
- Kalivas, P. W., Churchill, L., & Klitenick, M. A. (1993a). The circuitry mediating the translation of motivational stimuli into adaptive motor responses. In P. W. Kalivas & C. D. Barnes (Eds.), Limbic motor circuits and neuropsychiatry (pp. 237–287). Boca Raton, FL: CRC Press.
- Kalivas, P. W., Churchill, L., & Klitenick, M. A. (1993b). GABA and enkephalin projection from the nucleus accumbens and ventral pallidum to VTA. Neuroscience, 57, 1047–1060.
- Kalivas, P. W., Duffy, P., & Barrow, J. (1989). Regulation of the mesocorticolimbic dopamine system by glutamic acid receptor subtypes. Journal of Pharmacology and Experimental Therapeutics, 251, 378–387.
- Kalivas, P. W., Taylor, S., & Miller, J. S. (1985). Sensitization to repeated enkephalin administration into the ventral tegmental area of the rat: I. Behavioral characterization. Journal of Pharmacology and Experimental Therapeutics, 235, 537–543.
- Kalivas, P. W., Widerlov, E., Stanley, D., Breese, G. R., & Prange, A. J., Jr. (1983). Enkephalin action on the mesolimbic dopamine system: A dopamine-dependent and a dopamine-independent increase in locomotor activity. Journal of Pharmacology and Experimental Therapeutics, 227, 229–237.
- Kawaguchi, Y., Wilson, C. J., & Emson, P. C. (1990). Projection subtypes of the rat neostriatal matrix cells revealed by intracellular injection of biocytin. The Journal of Neuroscience, 10, 3421–3438.
- Kelley, A. E., & Domesick, V. B. (1982). The distribution of the projection from the hippocampal formation to the nucleus accumbens in the rat: An anterograde and retrograde horseradish peroxidase study. Neuroscience, 7, 2321–2335.
- Kim, J. H., & Vezina, P. (1997). Activation of metabotropic glutamate receptors in the rat nucleus accumbens increases locomotor activity in a dopamine-dependent manner. Journal of Pharmacology and Experimental Therapeutics, 283, 962–968.
- Kim, J. H., & Vezina, P. (1998). Metabotropic glutamate receptors are necessary for sensitization by amphetamine. NeuroReport, 9, 403–406.
- Kiyatkin, E. A. (1995). Functional significance of mesolimbic dopamine. Neuroscience and Behavioral Reviews, 19, 573–598.
- Kiyatkin, E. A., & Rebec, G. V. (1999). Striatal neuronal activity and responsiveness to dopamine and glutamate after selective blockade of D1 and D2 dopamine receptors in freely moving rats. The Journal of Neuroscience, 19, 3594–3609.
- Klitenick, M. A., Deutch, A. Y., Churchill, L., & Kalivas, P. W. (1992). Topography and functional role of dopaminergic projections from the ventral mesencephalic tegmentum to the ventral pallidum. Neuroscience, 50, 371–386.
- Klüver, H., & Bucy, P. C. (1939). Preliminary analysis of the functions of the temporal lobe in monkeys. Archives of Neurological Psychiatry, 42, 979–1000.
- Koob, G. F. (1992). Neuronal mechanisms of drug reinforcement. Annals of the New York Academy of Sciences, 654, 171–191.
- Kornetsky, C., & Espositio, R. U. (1979). Euphoriogenic drugs: Effects on reward pathways of the brain. Federation Proceedings, 38,
- Kuroda, M., Murakami, K., Shinkai, M., Ojima, H., & Kishi, K. (1995). Electron microscopic evidence that axon terminals from the mediodorsal thalamic nucleus make direct synaptic contacts with callosal cells in the prelimbic cortex of the rat. Brain Research, 677, 348–353.
- Lamour, Y., Dutar, P., Rascol, O., & Jobert, A. (1986). Basal forebrain neurons projecting to the rat frontoparietal cortex: Electrophysiological and pharmacological properties. Brain Research, 362,
- Lashley, K. S. (1938). Experimental analysis of instinctive behavior. Psychological Reviews, 45, 445–471.
- Le Moal, M., & Simon, H. (1991). Mesocorticolimbic dopaminergic network: Functional and regulatory roles. Physiological Review, 71, 155–234.
- Le Moine, C., & Bloch, B. (1995). D1 and D2 dopamine receptor gene expression in the rat striatum: Sensitive cRNA probes demonstrate prominent segregation of D1 and D2 mRNAs in the distinct neuronal population of dorsal and ventral striatum. The Journal of Comparative Neurology, 355, 418–426.
- Louilot, A., Le Moal, M., & Simon, H. (1986). Differential reactivity of dopaminergic neurons in the nucleus accumbens in response to different behavioral situations: An in vivo voltammetric study in freely moving rats. Brain Research, 397, 395– 400.
- Lu, X.-Y., Ghasemzadeh, M. B., & Kalivas, P. W. (1998). Expression of D1 receptor, D2 receptor, substance P and enkephalin messenger RNAs in the neurons projecting from the nucleus accumbens. Neuroscience, 82, 767–780.
- Maldonado-Irizarry, C. S., Swanson, C. J., & Kelley, A. E. (1995). Glutamate receptors in the nucleus accumbens shell control feeding behavior via the lateral hypothalamus. The Journal of Neuroscience, 15, 6779–6788.
- McBride, W. J., Murphy, J. M., & Ikemoto, S. (1999). Localization of brain reinforcement mechanisms: Intracranial selfadministration and intra-cranial place-conditioning studies. Behavioural Brain Research, 101, 129–152.
- McDonald, A. J. (1996). Glutamate and aspartate immunoreactive neurons of the rat basolateral amygdala: Colocalization of excitory amino acids and projections to the limbic circuit. The Journal of Comparative Neurology, 365, 367–379.
- McDonald, A. J., Mascagni, F., & Guo, L. (1996). Projections of the medial and lateral prefrontal cortices to the amygdala: A Phaseolus vulgaris leucoagglutinin study in the rat. Neuroscience, 71, 55–76.
- McFarland, K., & Ettenberg, A. (1995). Haloperidol differentially affects reinstatement and motivational processes in rats running an alley for intravenous heroin. Psychopharmacology, 122, 346– 350.
- McFarland, K., & Ettenberg, A. (1997). Reinstatement of drugseeking behavior produced by heroin-predictive environmental stimuli. Psychopharmacology, 131, 86–92.
- McFarland, K., & Ettenberg, A. (1998). Haloperidol does not affect motivational processes in an operant runway model of foodseeking behavior. Behavioral Neuroscience, 112, 630–635.
- McFarland, K., & Ettenberg, A. (1999). Haloperidol does not attenuate conditioned place preferences or locomotor activation produced by heroin- or food-predictive discriminative stimuli. Pharmacology, Biochemistry, and Behavior.
- McFarland, K., & Kalivas, P. W. (2001). Circuitry mediating cocaine-induced reinstatement. The Journal of Neuroscience.
- McGeer, P. L., Eccles, J. C., & McGeer, E. G. (1987). Molecular neurobiology of the mammalian brain. New York: Plenum
- Meil, W. M., & See, R. E. (1997). Lesions of the basolateral amygdala abolish the ability of drug associated cues to reinstate responding during withdrawal from self-administered cocaine. Behavioural Brain Research, 87, 139–148.
- Mogenson, G. J., Brudzynski, S. M., Wu, M., Yang, C. R., & Yim, C. C. Y. (1993). From motivation to action: A review of dopaminergic regulation of limbic-nucleus accumbenspedunculopontine nucleus circuitries involved in limbic-motor integration. In P. W. Kalivas & C. D. Barnes (Eds.), Limbic motor circuits and neuropsychiatry (pp. 193–236). Boca Raton, FL: CRC Press.
- Mogenson, G. J., Ciriello, J., Garland, J., & Wu, M. (1987). Ventral pallidum projections to mediodorsal nucleus of the thalamus: An anatomical and electrophysiological investigation. Brain Research, 404, 221–230.
- Mogenson, G. J., Jones, D. J., & Yim, C. Y. (1980). From motivation to action: Functional interface between the limbic system and the motor system. Progress in Neurobiology, 14, 69–97.
- Mogenson, G. J., & Nielson, M. (1984). A study of the contribution of hippocampal-accumbens-pallidal projections to locomotor activity. Behavioral and Neural Biology, 42, 38–51.
- Morris, R. G. M., Anderson, E., Lynch, G. S., & Baudry, M. (1986). Selective impariment of learning and blockade of long-term potentiation by an N-methyl-D-aspartate receptor antagonist, AP5. Nature, 319, 774–776.
- Mucha, R. F., & Herz, A. (1985). Motivational properties of kappa and mu opioid receptor agonists studied with place and taste preference conditioning. Psychopharmacology, 86, 274–280.
- Napier, T. C., & Mitrovic, I. (1999). Opioid modulation of ventral pallidal inputs. Annals of the New York Academy of Sciences, 877, 176–201.
- Napier, T. C., Mitrovic, I., Churchill, L., Klitenick, M. A., & Kalivas, P. W. (1995). Substance P projection from the nucleus accumbens to the ventral pallidum: Anatomy, electrophysiology and behavior. Neuroscience, 69, 59–70.
- Napier, T. C., Simson, P. E., & Givens, B. S. (1991). Dopamine electrophysiology of ventral pallidal/substantia innominata neurons: Comparison with the dorsal globus pallidus. Journal of Pharmacology and Experimental Therapeutics, 258, 249– 257.
- Nauta, W. J. H. (1961). Fiber degeneration following lesions of the amygdaloid complex in the monkey. Journal of Anatomy, 95, 515–531.
- Nisenbaum, E. S., Berger, T. W., & Grace, A. A. (1993). Depression of glutamatergic and GABAergic synaptic responses in striatal spiny neurons by stimulation of presynaptic GABAB receptors. Synapse, 14, 221–242.
- Nishikawa, T., Takasima, M., & Toru, M. (1983). Increased [3H]kainic acid-binding in the prefrontal cortex in schizophrenia. Neuroscience Letters, 40, 245–250.
- O’Donnell, P., & Grace, A. A. (1995). Synaptic interactions among excitatory afferents to nucleus accumbens neurons: Hippocampal gating of prefrontal cortical input. The Journal of Neuroscience, 15, 3622–3639.
- O’Donnell, P., & Grace, A. A. (1996). Dopaminergic reduction of excitability in nucleus accumbens neurons recorded in vitro. Neuropsychopharmacology, 15, 87–97.
- O’Donnell, P., Greene, J., Pabello, N., Lewis, B. L., & Grace, A. A. (1999). Modulation of cell firing in the nucleus accumbens. Annals of the New York Academy of Sciences, 877, 157–175.
- Olds, J., & Milner, P. (1954). Positive reinforcement produced by electrical stimulation of septal area and other regions of rat brain. The Journal of Comparative and Physiological Psychology, 47, 419–427.
- Olive, M. F., Anton, B., Micevych, P., Evans, C. J., & Maidment, N. T. (1997). Presynaptic versus postsynaptic localization of mu and delta opioid receptors in dorsal and ventral striatalpallidal pathways. The Journal of Neuroscience, 17, 7471–7479.
- Ono, T., Nishijo, H., & Nishino, H. (2000). Functional role of the limbic system and basal ganglia in motivated behavior. Journal of Neurology, 247(Suppl. 5), 23–32.
- Otake, K., & Nakamura, Y. (2000). Possible pathways through which neurons of the shell of the nucleus accumbens influence the outflow of the core of the nucleus accumbens. Brain Development, 22(Suppl. 1), 17–26.
- Parent, A., & Hazrati, L.-N. (1995). Functional anatomy of the basal ganglia: I. The cortico-basal ganglia-thalamo-cortical loop. Brain Research Reviews, 20, 91–128.
- Parkinson, J., Robbins, T., & Everitt, B. (2000). Dissociable roles of the central and basolateral amygdala in appetitive emotional learning. European Journal of Neuroscience, 12, 405–413.
- Pearl, S. M., & Glick, S. D. (1996). Prolonged antagonism of morphine-induced locomotor stimulation by kappa opioid agonists: Enhancement by prior morphine exposure. Neuroscience Letters, 212, 5–8.
- Percheron, G., Francois, C., Yelnik, J., & Fenelon, G. (Eds.). (1989). The primate nigro-striatal-pallido-nigral system: Not a mere loop. London: John Libbey.
- Pfeiffer, A., Brandt, V., & Herz, A. (1986). Psychotomimesis mediated by kappa opiate receptors. Science, 233, 774–776.
- Pornnarin, T., Floresco, S. B., & Phillips, A. G. (2000). Hyperlocomotion and increased dopamine efflux in the rat nucleus accumbens evoked by electrical stimulation of the ventral subiculum: Role of ionotropic glutamate and dopamine D1 Psychopharmacology, 151, 242–251.
- Reardon, F., & Mitrofanis, J. (2000). Organisation of the amygdalothalamic pathways in rats. Anatomy and Embryology, 201, 75–84.
- Robbins, T. W., Cador, M., Taylor, J. R., & Everitt, B. J. (1989). Limbic-striatal interactions in reward-related processes. Biobehavioral Reviews, 13, 155–162.
- Roberts, A. C., & Wallis, J. D. (2000). Inhibitory control and affective processing in the prefrontal cortex: Neurophysiological studies in the common marmoset. Cerebral Cortex, 10, 252– 262.
- Roberts, D. C. S., & Andrews, M. M. (1997). Baclofen suppression of cocaine self-administration: Demonstration using a discrete trials procedure. Psychopharmacology, 131, 271–278.
- Robinson, T., & Berridge, K. (1993). The neural basis of drug craving: An incentive-sensitization theory of addiction. Research Reviews, 18, 247–291.
- Rolls, E. T., Critchlet, H. D., Mason, R., & Wakeman, E. A. (1996). Orbitofrontal cortex neurons: Role in olfactory and visual association learning. Journal of Neurophysiology, 75, 1970–1981.
- Rosenkranz, J. A., & Grace, A. A. (1999). Modulation of basolateral amygdala neuronal firing and afferent drive by dopamine receptor activation in vivo. Neuroscience, 19, 11027–11039.
- Sacaan, A. I., Bymaster, F. P., & Schoepp, D. D. (1992). Metabotropic glutamate receptor activation produced extrapyramidal motor system activation that is mediated by striatal dopamine. Journal of Neurochemistry, 59, 245–251.
- Sala, M., Braida, D., Colombo, M., Groppetti, A., Sacco, S., Gori, E., et al. (1995). Behavioral and biochemical evidence of opioidergic involvement in cocaine sensitization. Journal of Pharmacology and Experimental Therapeutics, 274, 450–457.
- Salamone, J., Kurth, P., McCullough, L., Sokolowski, J., & Cousins, M. (1993). The role of brain dopamine in response initiation: Effects of haloperidol and regionally specific dopamine depletions on the local rate of instrumental responding. Brain Research, 628, 218–226.
- Salamone, J. D. (1994). The involvement of nucleus accumbens dopamine in appetitive and aversive motivation. Behavioural Brain Research, 61, 117–133.
- Schenk, S., Partridge, V., & Shippenberg, T. (2000). Reinstatement of exinguished drug-taking behavior in rats: Effect of the kappaopioid receptor agonist, U69593. Psychopharmacology, 151(1), 85–90.
- Schilstrom, B., Fagerquist, M., Zhang, X., Hertel, P., Panagis, G., Nomikos, G., et al. (2000). Putative role of presynaptic 7* nicotinic receptors in nicotine stimulated increases of extracellular levels of glutamate and aspartate in the ventral tegmental area. Synapse, 38, 413–420.
- Schneider, L. H., Davis, J. D., Watson, C. A., & Smith, G. P. (1990). Similar effects of raclopride and reduced sucrose concentration on the microstructure of sucrose sham feeding. European Journal of Pharmacology, 186, 61–70.
- Schultz, W. (1998). Predictive reward signal of dopamine neurons. American Journal of Physiology, 80, 1–27.
- Schultz, W., Apicella, P., & Ljungberg, T. (1993). Responses of monkey dopamine neurons to reward and conditioned stimuli during successive steps of learning a delayed response task. The Journal of Neuroscience, 13, 900–913.
- Seamans, J. K., Floresco, S. B., & Phillips, A. G. (1998). D1 receptor modulation of hippocampal-prefrontal cortical circuits integrating spatial memory with executive functions in the rat. The Journal of Neuroscience, 18, 1613–1621.
- Sesack, S. R., Deutch, A. Y., Roth, R. H., & Bunney, B. S. (1989). Topographical organization of the efferent projections of the medial prefrontal cortex in rat: An anterograde tract-tracing study with Phaseolus vulgais The Journal of Comparative Neurology, 290, 213–242.
- Sesack, S. R., & Pickel, V. M. (1990). In the rat medial nucleus accumbens, hippocampal and catecholaminergic terminals converge on spiny neurons and are in apposition to each other. Brain Research, 527, 266–272.
- Shreve, P. E., & Uretsky, N. J. (1989). AMPA, kalinic acid and N-methyl-D-aspartic acid stimulate locomotor activity after injection into the substantia innominata/lateral preoptic area. Pharmacology, Biochemistry, and Behavior, 34, 101– 106.
- Shreve, P. E., & Uretsky, N. J. (1990). GABAand glutamate interact in the substantia innominata/lateral preoptic area to modulate locomotor activity. Pharmacology, Biochemistry, and Behavior, 38, 385–388.
- Skinner, B. F. (1950). Are learning theories necessary? Psychological Reviews, 57, 193–216.
- Skinner, B. F. (Ed.). (1959). Animal research in the pharmacotherapy of mental disease. Washington, DC: National Academy of Sciences.
- Smiley, J. F., & Goldman-Rakic, P. S. (1993). Heterogeneous targets of dopamine synapses in monkey prefrontal cortex demonstrated by serial section electron microscopy: A laminar analysis using the silver-enhanced diaminobenzidine sulfide (SEDS) immunolabeling technique. Cerebral Cortex, 3(3), 223–238.
- Steiner, H., & Gerfen, C. R. (1998). Role of dynorphin and enkephalin in the regulation of striatal output pathways and behavior. Experimental Brain Research, 123, 60–76.
- Stellar, J. R., Kelley, A. E., & Corbett, D. (1983). Effects of peripheral and central dopamine blockade on lateral hypthalamic self-stimulation: Evidence for both reward and motor deficits. Pharmacology, Biochemistry, and Behavior, 18, 433–442.
- Stratford, T. R., & Kelley, A. E. (1997). GABA in the nucleus accumbens shell participates in the central regulation of feeding behavior. The Journal of Neuroscience, 17, 4434–4440.
- Stratford, T. R., Kelley, A. E., & Simansky, K. J. (1999). Blockade of GABAA receptors in the medial ventral pallidum elicits feeding in satiated rats. Brain Research, 825, 199–203.
- Swanson, L. W. (1982). The projections of the ventral tegmental area and adjacent regions: A combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Research Bulletin, 9, 321–353.
- Tamura, R., Ono, T., Fukuda, M., & Nishijo, H. (1992). Monkey hippocampal neuron responses to complex sensory stimulation during object discrimination. Hippocampus, 2, 287–306.
- Thomas, M., Malenka, R., & Bonci, A. (2000). Modulation of longterm depression by dopamine in the mesolimbic system. The Journal of Neuroscience, 20, 5581–5586.
- Timmerman, W., & Westerink, B. H. (1997). Brain microdialysis of GABAand glutamate: What does it signify? Synapse, 27, 242–261.
- Timofeev, I., Grenier, F., & Steriade, M. (1998). Spike-wave complexes and fast components of cortically generated seizures: IV. Paroxysmal fast runs in cortical and thalamic neurons. Journal of Neurophysiology, 80, 1495–1513.
- Tombaugh, T. N., Szostak, C., & Mills, P. (1983). Failure of pimozide to disrupt acquisition of light-dark and spatial discrimination problems. Psychopharmacology, 79, 161–168.
- Tremblay, L., & Schultz, W. (1999). Relative reward preference in primate orbitofrontal cortex. Nature, 382, 629–632.
- Uchimura, N., & North, R. A. (1991). Baclofen and adenosine inhibit synaptic potentials mediated by gamma-aminobutyric acid and glutamate release in the nucleus accumbens. Journal of Pharmacology and Experimental Therapeutics, 258, 663–668.
- Vezina, P., Kalivas, P. W., & Stewart, J. (1987). Sensitization occurs to the locomotor effects of morphine and the specific mu opioid receptor agonist, DAGO, administered repeatedly to the VTA but not to the nucleus accumbens. Brain Research, 417, 51–58.
- Vezina, P., & Kim, J. H. (1999). Metabotropic glutamate receptors and the generation of locomotor activity: Interactions with midbrain dopamine. Neuroscience and Biobehavioral Reviews, 23, 577–589.
- Walker, J. M., Thompson, L. A., Frascella, J., & Friederich, M. W. (1987). Opposite effects of mu and kappa opiates on the firingrate of dopamine cells in the substantia nigra of the rat. European Journal of Pharmacology, 134, 53–59.
- Watanabe, M. (1996). Reward expectancy in primate prefrontal neurons. Nature, 382, 629–632.
- Watson, J. B. (1913). Psychology as the behaviorist views it. Psychological Reviews, 20, 158–177.
- Watson, J. B. (1919). Psychology from the standpoint of a behaviorist. Philedelphia: J. B. Lippincott.
- Weiskrantz, L. (1956). Behavioral changes associated with the ablation of the amygdaloid complex in monkeys. The Journal of Comparative and Physiological Psychology, 49, 381–391.
- Willins, D. L., Narayanan, S., Wallace, L. J., & Uretsky, N. J. (1993). The role of dopamine and AMPA/kainate receptors in the nucleus accumbens in the hypermotility response to MK801. Pharmacology, Biochemistry, and Behavior, 46, 881–887.
- Willins, D. L., Wallace, L. J., Miller, D. D., & Uretsky, N. J. (1992). A-Amino-3-hydroxy-5-methylisoxazole-4-propionate/ kainate receptor antagonists in the nucleus accumbens and ventral pallidum decrease the hypermotility response to psychostimulant drugs. Journal of Pharmacology and Experimental Therapeutics, 260, 1145– 1151.
- Wilson, C. J., & Kawaguchi, Y. (1996). The origins of two-state spontaneous membrane potential fluctuations of neostriatal spiny neurons. The Journal of Neuroscience, 16, 2397–2410.
- Wise, R. A. (1978). Neuroleptic attenuation of intracranial selfstimulation: Reward or performance deficits? Life Science, 22, 535–542.
- Wise, R. A. (1982). Neuroleptics and operant behavior: The anhedonia hypothesis. Behavioral Brain Sciences, 5, 39–88.
- Wise, R. A., & Rompre, P. P. (1989). Brain dopamine and reward. Annual Review of Psychology, 40, 191–215.
- Wolf, M. E. (1998). The role of excitatory amino acids in behavioral sensitization to psychomotor stimulants. Progress in Neuorobiology, 54, 679–720.
- Woodworth, R. S. (1918). Dynamic psychology. New York: Columbia University Press.
- Wright, C. I., Beijer, V. J., & Groenewegen, H. J. (1996). Basal amygadaloid complex afferents to the rat nucleus accumbens are compartmentally organized. The Journal of Neuroscience, 16, 1877–1893.
- Xi, Z.-X., & Stein, E. (1999). Baclofen inhibits heroin selfadministration behavior and mesolimbic dopamine release. Journal of Pharmacology and Experimental Therapeutics, 290, 1369–1374.
- Xu, M., Moratalla, R., Gold, L. H., Hiroi, N., Koob, G. F., Graybiel, A. M., et al. (1994). Dopamine D1 receptor mutant mice are deficient in striatal expression of dynorphin and and in dopaminemediated behavioral responses. Cell, 79, 729–742.
- Yang, C. R., & Mogenson, G. J. (1985). An electrophysiological study of the neural projections from the hippocampus to the ventral pallidum and the subpallidal areas by way of the nucleus accumbens. Neuroscience, 15, 1015–1025.
- Yang, C. R., & Seamans, J. K. (1996). Dopamine D1 receptor actions in layers V-VI rat prefrontal cortex neurons in vitro: Modulation of dendritic-somatic signal integration. The Journal of Neuroscience, 16, 1922–1935.
- Yang, C. R., Seamans, J. K., & Gorelova, N. (1999). Developing a neuronal model for the pathophysiology of schizophrenia based on the nature of electrophysiological actions of dopamine in the prefrontal cortex. Neuropsychopharmacology, 21, 161– 194.
- Yeomans, J. S. (1990). Principles of brain stimulation. New York: Oxford University Press.
- Yim, C. Y., & Mogenson, G. J. (1988). Neuromodulatory action of dopamine in the nucleus accumbens: An in vivo intracellular study. Neuroscience, 26, 403–411.
- Yokel, R. A., & Wise, R. A. (1976). Attenuation of intravenous amphetamine reinforcement by central dopamine blockade in rats. Psychopharmacology, 48, 311–318.
- Zahm, D. S., & Brog, J. S. (1992). On the significance of subterritories in the “accumbens” part of the rat ventral striatum. Neuroscience, 50, 751–767.
- Zahm, D. S., & Heimer, L. (1990). Two transpallidal pathways originating in the rat nucleus accumbens. The Journal of Comparative Neurology, 302, 437–446.
- Zahm, D. S., Williams, E., & Wohltmann, C. (1996). Ventral striatopallidothalamic projection: IV. Relative involvements of neurochemically distinct subterritories in the ventral pallidum and adjacent parts of the rostroventral forebrain. The Journal of Comparative Neurology, 364, 340–362.
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality