
View sample genetic basis of personality structure research paper. Browse research paper examples for more inspiration. If you need a psychology research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our writing service for professional assistance. We offer high-quality assignments for reasonable rates.
Until recently, the study of personality was handicapped by the lack of a systematic taxonomy of constructs to represent individual differences. A confusing array of constructs and measures was available, and different measures of purportedly the same construct often showed little correspondence. This diversity hindered the development of a systematic understanding of individual differences. Recently, the situation began to change with emerging agreement about some of the major dimensions of personality. Broad traits such as neuroticism-stability, extraversion-introversion, and psychoticism-constraint are identified in most analyses of personality traits and part of most descriptive systems. There is also agreement about the way personality is organized. Models based on trait concepts assume that traits differ along a dimension of breadth or generalization and that traits are hierarchically organized, with global traits such as neuroticism subdividing into a set of more specific traits such as anxiousness and dependence (Goldberg, 1993; Hampson, John, & Goldberg, 1986).
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
Within this framework, attention has focused particularly on the five major factors as a parsimonious taxonomy of personality traits (Goldberg, 1990). Lexical analyses of the natural language of personality description (Digman, 1990; Goldberg, 1990) and subsequent psychometric studies of personality inventories (Costa & McCrae, 1992) have converged in identifying five broad factors typically labeled extraversion or surgency, agreeableness, conscientiousness, emotional stability versus neuroticism, and intellect, culture, or openness. It is widely assumed that this structure is transforming our understanding of personality and that the higherorder structure of personality is becoming more clearly delineated. Enthusiasm for the emergent structure, although understandable because it promises to bring coherence to a field characterized more by conceptual and theoretical debate than by substantive findings, tends to minimize confusions that still exist regarding the number and content of higherorder domains (Zuckerman, 1991, 1995, 1999; Zuckerman, Kulhman, Joireman, Teta, & Kraft, 1993) and nature of the assumed hierarchical arrangement of traits.
These problems remain unresolved despite numerous attempts to explicate personality structure, partly because the methods used incorporate subjective elements regarding choice of analytic strategies and data interpretation, and partly because personality concepts are inherently fuzzy, a factor that contributes to interpretive problems. In this research paper, we examine the contribution that behavioral genetic approaches can make to explicating the structure of personality and resolving issues of the number and content of domains. The argument we advance is that an approach that contributes to understanding of the causes of trait covariation (as opposed to approaches that simply offer descriptions of trait covariation) offer an important perspective on these intractable taxonomic problems.
Domain Definition: Unresolved Problems with Phenotypic Structure
Number of Domains
Despite the dominance of the five-factor approach, disagreement still exists on the number of dimensions required to represent the higher-order structure of personality. Almagor, Tellegen, and Waller (1995), for example, suggested that five factors do not capture all dimensions of the natural language of personality because lexical analyses excluded terms that were evaluative or described temporary states such as mood. When they used an unrestricted set of terms, seven factors were identified. Five factors—Positive Emotionality, Negative Emotionality, Dependability, Agreeableness, Negative Emotionality, and Conscientiousness—corresponded to the five-factor dimensions of Extraversion, Neuroticism, Conscientiousness, Agreeableness, and Openness (negatively), respectively. The remaining factors were evaluative dimensions, Positive Valence and Negative Valence, which are not represented in the five-factor model. They concluded that the seven-factor model provides a better representation of lexical descriptions of personality. McCrae and John (1992) and Widiger (1993) refuted this conclusion, claiming that positive and negative valence factors could be assumed under the five factors.
Whereas Almagor, Tellegen, and Waller (1995) maintained that the five-factor model is too parsimonious, Eysenck (1991) suggested that it is not parsimonious enough. He argued that the five domains differ in abstractness and that the five dimensions could be accommodated within his threefactor model of Psychoticism, Extraversion, and Neuroticism because the Openness andAgreeableness domains are merely facets of Psychoticism. Studies examining the relationship between NEO-PI-R (Neurosis Extraversion OpennessPersonality Inventory-Revised) and EPQ-R (Eysenck Personality Questionnaire-Revised), however, suggest that although the two scales overlap they assess unique aspects of personality (Avia et al., 1995; Draycott & Kline, 1995). These problems occur because the five factors, although assumed to be orthogonal, in fact intercorrelate. For example, correlations between NEO-PI-R Neuroticism and Conscientiousness domains and Extraversion and Openness to Experience domains are −.53 and .40 respectively (Costa & McCrae, 1992). These values raise the important issue of what degree of overlap or covariation between domains is tolerable. Whether these values are interpreted as unimportant or substantial depends largely on the investigator’s theoretical perspective.
Domain Definition
A related issue is lack of agreement on the lower-order traits that define each domain. Identification of an optimal set of lower-order traits has proved difficult (Costa & McCrae, 1998). Questions about whether a facet belongs to a proposed domain are raised when it consistently correlates with facets comprising another domain. For example, although Costa and McCrae (1992) report a moderate correlation of −.25 between total domain scores for Neuroticism and Agreeableness, the correlations between the Neuroticism facet Angry Hostility and Agreeableness facets Trust, Altruism, and Compliance are −.42, −.34, and −.49, respectively, and the correlation between Angry Hostility and the total Agreeableness domain score is −.47. Similarly, the correlation between the total Neuroticism domain score and the Agreeableness facet Trust is −.37. How this overlap is interpreted often forms the basis of many authors’claims as to why their model provides the “correct” description of personality. As with the intercorrelations among domains, the interpretations placed on the findings are largely arbitrary.
This problem is also revealed by factor analyses of facet scales. Although factor loadings may conform to simple structure and the hypothesized five-factor pattern, some facets may have an appreciably lower loading than do the other facets defining a domain. This occurs with the NEO-PI-R Neuroticism facet of Impulsiveness. The correlations between Impulsiveness and the other Neuroticism facets range from .31 to .40 (Costa & McCrae, 1992). The median intercorrelation is .35, whereas the median intercorrelation among the other facets is .57. Findings such as these raise questions about the definition of domains and the possibility that additional domains are required to provide a comprehensive taxonomy.
The issue of establishing a coherent set of traits for each domain is related to the interpretation of each domain. Even within the five-factor approach there are differences in the interpretation of some domains, especially the domain that Costa and McCrae label Openness to Experience. They emphasize such defining characteristics as artistic, curious, original, and having wide interests (McCrae & Costa, 1985a, 1985b). In the NEO-PI-R, the factor is defined by ideas (curious), fantasy (imagination), aesthetics (artistic), actions (wide interests), feelings (excitable), and values (unconventional). Others consider the domain to represent culture or intellect (Digman, 1990; Saucier & Goldberg, 1996). John and Srivastava (1999) maintained that the culture label (Passini & Norman, 1966) is not supported by evidence that traits referring to culture such as civilized, polished, dignified, foresighted, and logical load more highly on the conscientiousness factor. This leaves the alternative interpretation of intellect (Digman & Inouye, 1986; Goldberg, 1990). However, John and Srivastava (1999) concluded that the evidence supports the Costa and McCrae interpretation and that intellect is merely a component of a broader openness factor. This interpretation is supported by studies of the relationship between the domain and measures of cognitive ability. For example, the openness-intellect factor (Understanding, Sentience, Change, and Autonomy) based on the Personality Research Form (Jackson, 1984) correlates highly with measures of crystallized intelligence (e.g., verbal subscales) but less with measures of fluid ability (arithmetic and performance subscales; Ashton, Lee, Vernon, & Jang, 1999).
There are other, less easily resolved confusions about the definition and facet structure of other domains. For example, Conscientiousness according to Costa and McCrae (1995) consists of a single factor defined by competence, order, dutifulness, achievement striving, self-discipline, and deliberateness. Paunonen and Jackson (1996), however, question the unity of conscientiousness: “. . . The domain is best thought of as three separate, but somewhat overlapping, dimensions related to being (a) methodical and orderly, (b) dependable and reliable, and (c) ambitious and driven to succeed. Moreover, the amount of overlap among these three facets may not be high enough to justify their inclusion in an overall Conscientiousness measure” (p. 55).
The cluster of traits labeled impulsive–sensation seeking poses an even greater problem. Earlier, we discussed problems with the placement of impulsiveness within the fivefactor model. The controversy, however, is deeper. For Zuckerman (1991, 1994), impulsivity and sensation seeking define a separate higher-order factor within an alternative five-factor structure. The factor resembles Eysenck’s psychoticism and Tellegen’s (1985) constraint. There appears, therefore, to be strong support for this domain. The five-factor model of Costa and McCrae, however, divides this factor into impulsivity and sensation seeking and assigns them to different domains. Impulsivity is considered part of neuroticism, an interpretation that is not shared by other conceptions of neuroticism, whereas sensation seeking is assigned to extraversion. This leads to similar problems with extraversion.
Extraversion is defined by subsets of traits that differ across models. These subsets include such traits as sociability or affiliation, agency, activation, impulsive–sensation seeking, positive emotions, and optimism (Depue & Collins, 1999; Watson & Clark, 1997). Depue and Collins (1999) pointed out that most accounts of extraversion postulate two central features, an interpersonal engagement component consisting of affiliation or sociability and agency, and an impulsivity component that includes sensation seeking. They suggested that impulsive–sensation seeking arises from the interaction of extraversion and a second independent trait represented by Tellegen’s (1985) constraint. This proposal differs, however, from Eysenck’s model that places impulsivity in the psychoticism domain and Costa and McCrae’s proposal that it belongs to neuroticism. It also differs from Gray’s (1973, 1987; Pickering & Gray, 1999) model that considers impulsivity as assessed by questionnaire to be a blend of Eysenck’s higher-order dimensions of extraversion and psychoticism. It appears, therefore, that there are major unresolved definitional problems with most domains that compromise claims that the five-factor model provides a basic assessment framework (McCrae & Costa, 1986).
The existence of such basic uncertainty about the taxonomy of personality traits would seem to suggest that statements that the structure of personality is becoming delineated might be a little premature. Uncertainty about the relationships among traits is a major obstacle to constructing a theory of individual differences and clarification of these issues is essential for the field to advance. The ordering of traits within each domain forms the basis for developing theoretical explanations by defining relationships that require explanation. In effect, a descriptive taxonomy shapes subsequent research and theory development.
Approaches to Domain Definition
In response to these challenges, especially Paunonen and Jackson’s (1996) critique of conscientiousness, Costa and McCrae (1998) outlined six methodological approaches that can be used to demonstrate the unity of any domain: (a) item content analysis, (b) definitions of psychological opposites, (c) examination of empirical correlates, (d) interpreting secondary and tertiary factor loadings, (e) identification of equivalents in specialized languages and (f) case studies. Costa and McCrae (1998) applied these approaches to show that the Conscientiousness domain was unitary in nature. The limitation of these proposals is their reliance on an array of criteria that incorporate a subjective element. The proposal relies on a convergence of evidence across sets of traditional phenotypic and psychometric analyses. However, numerous psychometric studies have not resolved these problems, raising the possibility that studies of phenotypes alone may not be sufficient.
The problem with phenotypic analyses is their reliance on constructs that are by their nature fuzzy and imprecise. This is illustrated by the confusion noted about the components of extraversion (Depue & Collins, 1999; Watson & Clark, 1997). Conceptions of extraversion include sociability or affiliation (includes agreeableness, affiliation, social recognition, gregariousness, warmth, and social closeness), agency (surgency, assertion, endurance, persistence, achievement, social dominance, ascendancy, ambitiousness), activation (liveliness, talkativeness, energy level, activity level, activity level), impulsive–sensation seeking (impulsivity, sensation seeking, excitement seeking, novelty seeking, boldness, risk taking, unreliability, disorderliness, adventurousness, thrill and adventure seeking, monotony avoidance, boredom susceptibility), positive emotions (positive affect, elatedness, enthusiasm, exuberance, cheerfulness, merriness, joviality), and optimism (Depue & Collins, 1999).
This list reveals the problems faced by attempts to delineate phenotypic structure. Not only does the content of extraversion differ across models, but the definition of each basic or lower-order trait may also differ across models and measures. Moreover, the meaning of putatively distinct traits overlaps so that facet traits defining a given domain shade into each other and into facet traits defining other domains. This fuzziness is probably an inevitable consequence of using natural language concepts that evolved to capture socially significant behaviors that are multidetermined. It adds to concerns that the taxonomies of phenotypic traits may not represent natural cleavages in the way behavior is organized nor reflect underlying etiological structures.
This fuzziness contributes to the considerable variability in personality phenotypes so that minor variations in measures and samples influence the number and contents of factors. The problem is compounded by the fact that many decisions about methodology and analytic strategies have an arbitrary component. More objective criteria are needed to guide decisions on the number of higher-order domains and the location of lower-order or basic traits within domains and to define a systematic set of basic traits. Phenotypic analyses are concerned primarily with describing trait covariation. This evokes the oft-voiced criticism of the five-factor approach—it is descriptive rather than explanatory. The basic problem of why traits are related to each other is not considered. An understanding of etiology of trait covariance, especially genetic etiology, would provide a conceptual foundation for current models. At each level of the trait hierarchy, traits and behaviors, including test items, could be grouped according to a shared etiology. Etiology would provide an additional criterion to supplement the usual psychometric criteria such as proposed by Costa and McCrae (1997) to guide decisions on the number and content of domains. Identification of a robust model of personality structure would be facilitated by evidence that a given phenotypic structure reflects the genetic architecture of personality traits. Unfortunately there are few studies of the genetic architecture underlying multiple personality traits compared to studies of phenotypic structure. Evidence that a given phenotypic structure parallels genotypic structure would support the validity and generalizability of the structure.
Heritability
The foundation for an etiological understanding of personality structure and for a behavioral genetic approach is provided by evidence that genetic influences account for approximately 40–60% of the variance for virtually all personality traits, with most of the remaining variance being explained by nonshared environmental effects (Bouchard, 1999; Loehlin & Nicholls, 1976; Plomin, Chipeur, & Loehlin, 1990). The broad traits of extraversion and neuroticism have received most attention. The data from several twin studies yield heritability estimates of approximately 60% for extraversion and 50% for neuroticism. Loehlin (1992) also examined multiple personality scales organized according to the five-factor framework. Estimates of about 40% heritability were obtained for each domain. Subsequent studies using the NEO-PI-R yielded heritability estimates of 41% for neuroticism, 53% for extraversion, 41% for agreeableness, and 40% for conscientiousness (Jang, Livesley, Vernon, & Jackson, 1996; see also Bergeman et al., 1993; Jang, McCrae, Angleitner, Riemann, & Livesley, 1998). Nonadditive genetic effects accounted for 61% the variance in openness to experience.
Although the evidence points to a significant genetic component to personality traits, it has been suggested that traits could be divided into temperament traits that have a substantial heritable component and character traits that are largely environmental in origin. If this is the case and environmental factors give rise to distinct traits, the role of genetic criteria in clarifying trait structure would be limited. The evidence does not, however, support the proposal. Putatively characterological traits such as openness to experience are as heritable as so-called temperament traits. Moreover, molecular genetic studies have found significant allelic associations between so-called character traits such as cooperativeness and self-directedness as assessed using the Temperament and Character Inventory and the 5-HTTLPR allele (Hamer, Greenberg, Sabol, & Murphy, 1999).
To date, a self-report measure of personality that has no genetic influence has not been identified (Plomin & Caspi, 1998).Thequalificationshouldbeaddedthatheritabilitystudies have relied largely on self-report measures—alternative methods of assessment may yield different results. However, this was not the case with the few studies using other methods (Heath, Neale, Kessler, Eaves, & Kendler, 1992; Riemann, Angleitner, & Strelau, 1997). Riemann and colleagues (1997), for example, reported a twin study conducted in Germany and Poland that compared assessments of the five factors using self-report questionnaires with peer ratings. Estimates of heritability based on self-report were similar to those reported by other studies. The peer ratings also showed evidence of heritability, although estimates were lower than those obtained from self-reports. Multivariate genetic analyses showed that the same genetic factors contributed to self-report and peer ratings. These results suggest that findings of a heritable component to all self-report measures are likely to generalize to other methods of measurement.
Evidence of heritability alone, however, is not sufficient to justify the use of behavioral genetic criteria to clarify trait structure. It is possible that environmental factors that account for about 50% of the variance have a substantial effect on trait covariation. If this were the case, the finding that traits are genetically related would be of less value in clarifying personality structure. The evidence, however, suggests that the phenotypic structure of traits closely parallels the underlying genetic architecture (Livesley, Jang, & Vernon, 1998; Loehlin, 1987)—a point that is discussed in detail later in this research paper.
It should be noted, however, that information about heritability merely explains the variance in a single trait as opposed to the covariance between traits. Such information has limited value in explicating personality structure. As Turkheimer (1998) argued, all individual differences in behavior are heritable and “. . . the very ubiquity of these findings make them a poor basis for reformulating scientists’ conceptions of human behavior” (p. 782). Nevertheless, information on heritability forms the foundation for understanding of the etiology of personality. The major contribution of behavior genetics to understanding personality structure, however, comes from multivariate genetic analyses that elucidate the genetic structure underlying multiple traits (Carey & DiLalla, 1994). Multivariate analyses extend univariate analysis of the genetic and environmental influences on a trait to evaluate genetic and environmental components of the covariation between two or more traits (DeFries & Fulker, 1986). It is this extension that promises to contribute to personality theory by explicating the etiological basis for trait covariance by evaluating the degree to which different traits are influenced by the same genetic and environmental factors. This issue is central to resolving some of the problems of personality description and structure.
The Etiological Basis of Covariance
The phenotypic covariation between two traits may be due to pleiotropy—that is, the degree to which the traits share a common genetic influence, environmental effects common to both traits, or both. The degree to which two variables have genetic and environmental effects in common is indexed by genetic (rG) and environmental correlation coefficients (rE). These statistics are interpreted as any other correlation coefficient and they may be subjected to other statistical procedures such as factor analysis (Crawford & DeFries, 1978). Genetic and environmental correlation coefficients are readily estimated from data obtained from monozygotic (MZ) and dizygotic (DZ) twin pairs.
The calculation of the genetic correlation is similar to that used to estimate the heritability of a single variable. A higher within-pair correlation for MZ twins than for DZ twins suggests the presence of genetic influences because the greater similarity is directly attributable to the twofold increase in genetic similarity in MZ versus DZ twins. In the multivariate case, a common genetic influence is suggested when the MZ cross-correlation (the correlation between one twin’s score on one of the variables and the other twin’s score on the other variable) exceeds the DZ cross-correlation. The phenotypic correlation (rp) between two variables (traits), x and y, is expressed by the following equation:
rp = (hx · hy · rg) + (ex · ey · re) (3.1)
where the observed or phenotypic correlation, (rp), is the sum of the extent to which the same genetic (rg) and/or environmental factors (re) influence each variable, weighted by the overall influence of genetic and environmental causes on each variable (hx, hy, ex, ey, respectively). The terms h and e are the square roots of heritability and environmental effect (h2 and e2) for variables x and y, respectively.
It should be noted that a genetic correlation describes statistical pleiotropism—that is, the extent to which allelic effects on trait predict allelic effects on the other trait. As Carey (1987) pointed out, statistical pleiotropism is not to be confused with biological pleiotropism in which two variables share the same loci. Unlike statistical pleiotropism, biological pleiotropism unequivocally links actual genes to behavior.
Phenotypic Structure and Genetic Architecture of Personality
A critical issue for understanding the etiological structure of personality and for the use of multivariate genetic analyses to clarify personality structure is the degree to which the phenotypic organization of traits reflects an underlying biological structure as opposed to the influence of environmental factors. The evidence indicates that the phenotypic structure of traits closely resembles the underlying genetic architecture and to a lesser degree environmental structure. The evidence also suggests that environmental factors do not appreciably influence trait covariation. These conclusions are based on comparisons of the factors extracted from matrices of phenotypic, genetic, and environmental correlations computed among traits comprising a given model or measure.
In one of the earliest studies of this kind, Loehlin (1987) analyzed the structure of item clusters from the California Psychological Inventory (CPI; Gough, 1989) in samples of MZ and DZ twins. Three matrices were derived that represented the covariance among different traits due to genetic, shared environmental, and nonshared environmental factors. When these matrices were examined with factor analysis, four factors emerged from analyses of genetic covariance that could be interpreted as representing Neuroticism, Extraversion, Openness, and Conscientiousness (few items related to the fifth factor, Agreeableness, are included in the CPI; see McCrae, Costa, & Piedmont, 1993). Analysis of shared environmental effects yielded two factors: family problems and masculinity-femininity. The former is not an aspect of personality per se, and the latter is probably an artifact of the exclusive use of same-sex twins (Loehlin, 1987). It should be noted, however, that shared environmental effects make relatively little contribution to the variance of personality traits. Hence, the important finding is the structure of nonshared environmental effects. Analysis of the nonshared environmental covariance matrix yielded three interpretable factors that resembled Neuroticism, Extraversion, and Conscientiousness. Thus, the structure of nonshared environmental influences largely mirrored genetic influences. This is not an isolated finding: Livesley et al. (1998) found similar structures in genetic and nonshared environmental components of traits related to personality disorder.
Livesley and colleagues (1998) examined the congruence of genetic and phenotypic factor structures and compared phenotypic structure across samples of personality disordered patients and two samples recruited from the general population. The clinical sample consisted of 602 patients with personality disorder. The general population samples consisted of 939 volunteer general population participants and 686 twin pairs. The twin sample allowed the computation of matrices of genetic and environmental correlations that could be compared against the phenotypic structures from all three samples. Personality was assessed with the Dimensional Assessment of Personality Pathology (DAPP; Livesley & Jackson, in press). This measure assesses 18 traits underlying personality disorder diagnoses that were identified in previous studies using a combination of clinical judgments, rational methods, and psychometric procedures (Livesley, 1986; Livesley, Jackson, & Schroeder, 1992).
Phenotypic correlations were computed in all three samples separately, and genetic and environmental correlations were computed on the twin sample. The phenotypic, genetic, and environmental correlation matrices were subjected to separate principal components analyses with rotation to oblimin criteria. Phenotypic structure was similar across all samples. Four factors were extracted from all five matrices (see Tables 3.1 and 3.2).
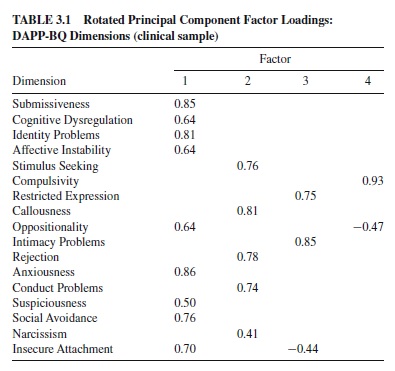
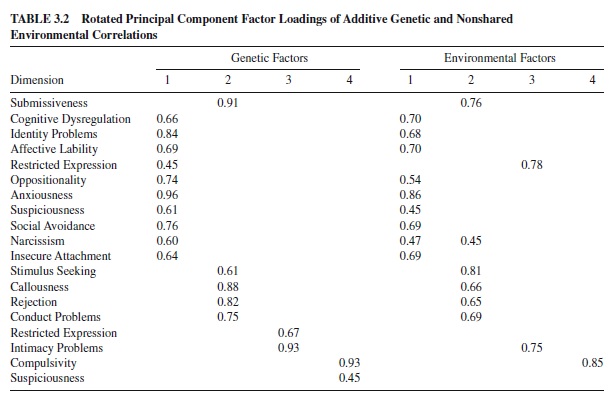
The first factor, Emotional Dysregulation, represents unstable and reactive affects and interpersonal problems. The factor resembled neuroticism as measured by the NEO-PI-R (Costa & McCrae, 1992; Schroeder, Wormworth, & Livesley, 1992)ortheEysenckPersonalityQuestionnaire(EPQ;Jang& Livesley, 1999) and the DSM-IV diagnosis of borderline personality disorder. The second factor, Dissocial Behavior, was negatively correlated with NEO-PI-R Agreeableness. It described antisocial traits and resembled the DSM-IV Cluster B antisocial personality diagnosis, Eysenck’s Psychoticism, and Zuckerman’s Impulsive–Sensation Seeking. The third factor, labeled Inhibition, was defined by intimacy problems and restricted expression of inner experiences and feelings. The factor correlated negatively with NEO-PI-R and EPQ Extraversion and resembled the DSM-IV avoidant and schizoid personality disorders. The fourth factor, Compulsivity clearly resembled NEO-PI-R Conscientiousness and DSM-IV obsessive-compulsive personality disorder. The loadings derived from the phenotypic correlation matrices were remarkably similar: Congruence coefficients ranged from .94 to .99. The congruency coefficients between the genetic and phenotypic factors on Emotional Dysregulation, Dissocial, Inhibition,andCompulsivitywere.97,.97,.98,and.95,respectively. The congruence between factors extracted from the phenotypic and nonshared environmental matrices were also high at .99, .96, .99, and .96, respectively. These data suggest that the phenotypic structure of personality and personality disorder traits closely reflects the underlying etiological architecture.
This conclusion is also supported by a study of the phenotypic structure and genetic architecture of the five-factor model assessed using the NEO-PI-R in two independent samples of twins recruited in Germany and Canada (Jang, Livesley, Angleitner, Riemann, & Vernon, in press). Factor analysis of the genetic and nonshared environmental covariance matrices yielded five factors that strongly resembled N, E, O,A, and C (Neurotic, Extraversion Openness,Agreeableness, and Conscientiousness). Congruence coefficients computed between the genetic factors and the published normative structure were .83, .72, .92, .88, and .70 for N, E, O, A, and C, respectively. The congruence of the nonshared environmental factors and normative structure was even higher at .96, .93, .90, .93, and .97 for N, E, O, A, and C, respectively.
The interesting feature of these results is not only that phenotypic structure resembles genetic structure, but also that the structure of environmental effects is similar to the genetic structure. Plomin, DeFries, and McClearn (1990) noted that across a range of studies, “the structure of genetic influences seems to be similar to the structure of [nonshared] environmental influences” (p. 236).They added that this is surprising: “Most of us would probably predict different patterns of genetic and environmental influences” (p. 236). Recently, however, it has been suggested that genetic factors are more important than are environmental influences in shaping trait structure because the resemblance of the structure of nonshared environmental effects to the observed structure of traits may be artifactual (McCrae, Jang, Livesley, Riemann, & Angleitner, in press).
Nonshared environmental effects are usually estimated as a residual term that may include systematic bias such as that introduced by implicit personality theory. Passini and Norman (1966) demonstrated this bias by asking students to rate the personalities of complete strangers. Although each rating was presumably a guess, a clear pattern to the ratings was found. Students who assumed that strangers were talkative also assumed that they were sociable and cheerful.Across a range of targets, these associations defined an Extraversion factor. Factor analysis of the ratings yielded the familiar five factors. Some researchers concluded from such studies that trait structure merely reflects the effects of semantic biases on person perceptions (Shweder, 1975). Ratings of strangers must contain bias due to implicit personality theory because they cannot be influenced by the true personalities of the targets. It is also likely that self-reports and ratings of wellknown targets incorporate a similar bias. For example, two observers may agree that a person is sociable but disagree on the extent of his or her sociability. The observer assigned a higher rating for sociability is also likely to assign a higher rating for cheerfulness and talkativeness. Thus, part of the covariance of these traits may be attributable to systematic biases in person perception that lead to correlated errors in individual judgments. If this is the case, similarities in structure between genetic covariance and nonshared environmental covariance could reflect the biasing effects of implicit personality theory on the latter.
To test for this bias, self-report twin data were supplemented with cross-observer correlations on the NEO-PI-R. This allowed the computation of two matrices of nonshared environmental covariance. The first estimated the covariance due to implicit personality theory bias alone. Factorial analysis of this matrix yielded the familiar five factors. Comparison with normative structure yielded congruence coefficients of .81, .45, .81, .89, and .85 for Neuroticism, Extraversion, Openness, Agreeableness, and Conscientiousness, respectively. The second matrix of nonshared environmental covariance estimated was free from systematic bias. Factor analysis of this “unbiased” matrix with targeted rotations to the normative NEO-PI-R factors produced low congruence coefficients at .53, .68, .22, .61, and .80 for Neuroticism, Extraversion, Openness, Agreeableness, and Conscientiousness, respectively. Subsequent factor analysis of this matrix yielded two factors. The first resembled a broad form of Conscientiousness with salient loading of the facets Activity, Order, Dutifulness, Achievement Striving, Self-Discipline, and (low) Impulsiveness. The second factor was defined by the facetsWarmth, Gregariousness, Positive Emotions, Openness to Feelings, Altruism, and Tender-Mindedness. This combination of Extraversion andAgreeableness facets resembles the Love axis of the Interpersonal Circumplex (Wiggins, 1979). The other interpersonal axis—Dominance—does not appear to be influenced by the nonshared environment. Assertiveness did not load on either factor.
These results suggest that when the conventional estimates of nonshared environmental covariances are decomposed into implicit personality theory bias and true nonshared effects, much of the resemblance to the five-factor structure appears attributable to bias. Overall, these studies point to the conclusion that genetic factors are largely responsible for the observed pattern of trait covariation.
The Hierarchical Structure of Personality
Beyond problems with the content of personality taxonomies, there are also uncertainties about the nature of the proposed hierarchical structure of traits and the relationship between higher- and lower-order traits. Factor analytic studies provide consistent evidence that specific traits are organized into more global entities. Lexical studies also show that natural language reflects this structure. Substantial agreement exists among individuals in judgments of trait breadth (Hampson et al., 1986). Despite this evidence, the nature and origins of the hierarchy are unclear. This is clearly a problem that requires explanation.
Fundamental differences exist among models on the way the personality hierarchy is conceptualized. The lexical approach seems to consider the higher-order domains to be lexical categories that impose structure on personality descriptors by organizing them into clusters that are not necessarily discrete or equally important (Saucier & Goldberg, 1996). The lexical structure “provides a framework for description, but not necessarily for explanation” (Saucier & Goldberg, 1996, p. 24–25). Saucier and Goldberg also asserted that “as a representation of phenotypes based on natural language, the Big Five structure is indifferent and thus complementary to genotypic representations of causes, motivations, and internal personality dynamics” (p. 42). The higher-order terms do not appear, therefore, to have any significance beyond that of description.
Traits psychologists, including other five-factor theorists, make different assumptions. For Allport (1961), a trait is “a neuropsychic structure” (p. 347) and therefore an explanatory concept. Eysenck also adopted this approach: Traits have heritable biological basis. Similarly, the five-factor model assumes that traits are “endogenous basic tendencies” with a substantial heritable component (McCrae & Costa, 1996, p. 72). For Eysenck and Costa and for McCrae, traits are explanatory as well as descriptive. In contrast to the lexical approach, the five-factor model assumes that domains are equally important and equal in breadth.
Assumptions that trait theories make about the psychobiological basis for the higher-order domains initially created uncertainty about the status of the lower-order traits. Most research effort has been directed toward understanding higher-order factors and little attention has been paid to parsing these domains into more specific components. Until recently, it was unclear whether the lower-order traits were merely facets of the higher-order traits or distinct entities with their own etiology. The use of the term facet to describe the lower-order traits, a convention adopted by Costa and McCrae, implies that they are merely exemplars or components of a more fundamental global trait. In this sense, the facet traits can be understood in terms of the domain sampling approach used in test construction in which facets are merely arbitrary ways to subdivide global traits to ensure adequate domain sampling. Identification of general genetic factors that have a broad influence on personality phenotypes also raises questions about the significance of the lower-order or facet traits—in particular, whether these traits are heritable simply because of their association with the broader domains or whether they are also subject to specific genetic influences. Clarification of this issue is critical to constructing an explanatory account of personality structure.
Heritability of Lower-Order Traits
If lower-order traits are only subcomponents of broader traits, all variance in a facet apart from error variance should be explained by the variance in the global trait. Recently, however, behavioral genetic research has suggested that lower-order traits have a distinct heritable component (Jang et al., 1998; Livesley et al., 1998). These studies estimated whether lowerorder traits have a unique genetic basis when the heritable component of higher-order traits is removed from them. Jang and colleagues (1998) partialled out all of the common variance due to each of higher-order Neuroticism, Extraversion, Openness,Agreeableness, and Conscientiousness scales from the 30 facet scales of the NEO-PI-R. When the residual variances on the facets were subjected to heritability analyses, a substantial genetic influence remained. Additive genetic effects accounted for 25 to 65% of the reliable specific variance, with most heritabilities ranging from .20 to .35 (see Table 3.3).
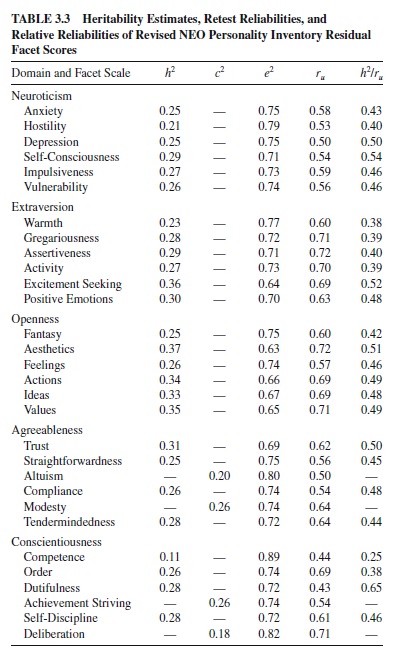
When these values were corrected for unreliability, the values increased to the usual range observed for personality traits. The implication is that these traits are not merely facets of more general traits, but rather distinct heritable entities.
A similar approach was used to study the residual heritability of the 18 traits underlying personality disorder (Livesley et al., 1998). Factor scores were computed for the four factors described previously. A standardized residual score for each scale was computed by regressing the four factor scores on each of the 18 basic traits. Monozygotic twin correlations were higher that the dizygotic twin correlations for all 18 traits. Estimates of the heritability of the residual trait scores showed substantial residual heritability for 11 of the 18 basic traits that ranged from .26 for Intimacy Problems to .48 for Conduct Problems.
These studies, in contrast to studies of phenotypic structure, point to the significance of the lower-order traits. Although these traits have tended to be neglected in personality research, they appear to be important for understanding personality. This suggests that a bottom-up approach to personality structure would provide additional information to complement that provided by the traditional top-down approach of the three- and five-factor models that identify the higher-order domains first and then seek to define an appropriate complement of facet traits. Before considering these issues in greater depth, it is important to recognize a limitation of the methods used. The regression method does not model genetic effects directly, and the results need to be replicated using multivariate genetic analyses. This introduces another feature of behavioral genetic analyses that is pertinent to understanding the genetic basis of personality: the use of pathways models to evaluate competing models of personality.
Independent and Common Pathways Models
In heritability analyses, components of variance are estimated by fitting models to the observed covariance matrices. In the univariate case, the heritability of a variable is estimated by comparing the similarity (estimated by Pearson’s r) of MZ to DZ twins. In the bivariate case, common genetic influences are suggested when the MZ cross-correlation exceeds the DZ cross-correlation used to compute the genetic correlation, rG. The multivariate extension of this idea is found in two general classes of path analytic models that are pertinent to personality research: independent and common pathways models (see Figures 3.1 and 3.2; McArdle & Goldsmith, 1990; Neale & Cardon, 1992). The independent pathway model specifies direct links from one or more genetic and environmental influences common to each variable and unique genetic and environmental effects to each variable. The common pathways model is a more stringent version of the independent pathways model. The primary difference between the two models is that the common pathways model postulates that of the covariation in a set of variables is mediated by a single latent variable that has its own genetic and environmental basis. Both models provide the opportunity to examine variance specific to each variable— that is, each lower-order trait. Factor analytic studies of personality have been concerned with reducing the covariance between lower-order traits to fewer factors. Residual variance specific to each trait is neglected. Biometric path models applied to twin data decompose this variance into etiological components. This makes it possible to evaluate the significance of these specific traits.
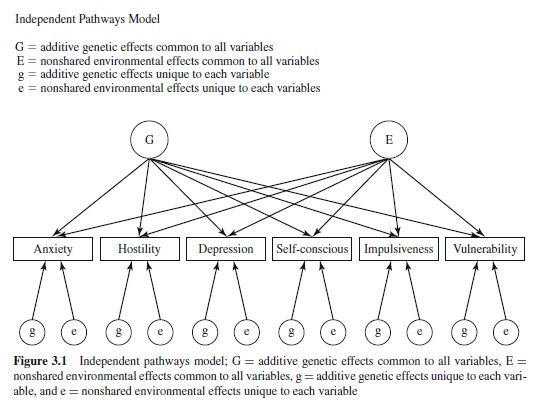
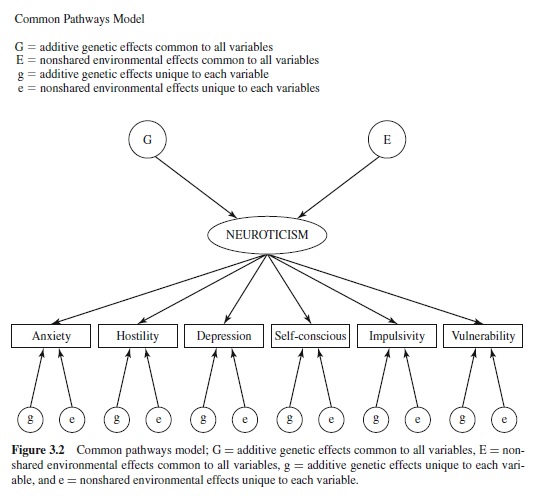
These models offer the opportunity to evaluate the hierarchical structure of personality by comparing the fit of the two models to the same data set. The common pathways model is the biometric equivalent to the traditional model of exploratory factor analysis used to delineate the phenotypic structure of traits. As applied to each of the five-factor domains, the model postulates a single latent factor for each domain that mediates the effects of genetic and environmental effects on each lower-order trait. In the case of NEO-PI-R Neuroticism, a latent variable of neuroticism is hypothesized through which genetic and environmental factors influence the six facets of Anxiety, Hostility, Depression, SelfConsciousness, Impulsivity, and Vulnerability. In contrast, the independent pathways model postulates direct genetic and environmental effects on each facet trait. The fits of these models provide an opportunity to evaluate different conceptions of personality structure. If the common pathways model provides the best fit, the implication is that the hierarchical structure of personality arises from the effects of higher-order factors that have a genetic and environmental basis. The task is then to explain how this entity differs from lower-order or facet traits and the role it plays in the formation of the hierarchy. If the independent pathways model provides the best fit, however, the implication is that the higher-order constructs of phenotypic analyses do not reflect the effects of a phenotypic entity, but rather the pleiotropic action of the genes shared by all lower-order or facet traits that define the domain. Under these circumstances, the task is to explicate the mechanisms that lead to trait clusters. Regardless of which model provides the best fit to the data, a useful feature of both models is that the magnitude of the path coefficients between each facet scale and the common genetic factor or latent variable along with information on the magnitude of genetic and environmental influences unique to each facet provides the basis for determining which facets should be grouped together within the taxonomy.
Five-Factor Model
Jang and colleagues (in press) fit common and independent pathways models to evaluate the coherence of the five domains assessed with the NEO-PI-R. The models were applied separately to a sample of 253 identical and 207 fraternal twin pairs from Canada and 526 identical and 269 fraternal pairs from Germany. The two samples made it possible to examine the universality of the etiological basis for personality structure by investigating whether the same genetic and environmental factors influenced personality traits in the two samples and whether they had similar effects.
For each sample, a single-factor common pathways model and a series of independent pathway models specifying variable numbers of genetic and nonshared environmental factor were fit to the six facets defining each domain. Shared environmental effects were omitted from the models because their effects were minimal. For each domain, the best fit was obtained with an independent pathways model. Table 3.4 illustrates the findings for the Neuroticism domain. An independent pathways model specifying two genetic factors and two nonshared environmental factors provided the most satisfactory explanation of the covariance between the six Neuroticism facets in the two samples. In both samples, the first genetic factor was marked by the Angry Hostility facet and, to a lesser extent, Anxiety. The second factor influenced all facets except Angry Hostility and Impulsivity. The depression facet had the highest loading in both samples.

In addition to demonstrating that the independent pathways model provided the best fit, these findings also suggest that the broad domains of personality are nonhomogeneous. This raises important questions about the factors that account for the apparent hierarchical structure of personality traits and the nature and conceptual status of the higher-order dimensions. These conclusions are, however, based on a single study using only a single measure of personality. Replication is clearly needed, given the results’ significance for understanding trait structure. The conclusions are, however, similar to those drawn from a study of personality disorder traits (Livesley & Jang, 2000).
Personality Disorder Traits
Livesley and Jang (2000) investigated the etiological structure of personality disorder by fitting independent and common pathways models to the 18 lower-order traits of personality disorder assessed by administering the DAPP to a volunteer sample of 686 twin pairs. Each trait consists of two or more specific traits so that a total of 69 specific traits define the 18 basic traits. The 18 traits in turn define four higherorder factors. Thus the DAPP system incorporates three levels of construct (higher-order factors, lower-order traits, and specific traits) whereas the NEO-PI-R has only two levels (domains and facets). This makes it possible to explore the genetic architecture of personality in more detail. For example, the basic trait of Anxiousness is defined by four specific traits: trait anxiety, guilt proneness, rumination, and indecisiveness. Each basic trait represents a single phenotypic factor. If personality is inherited as a few genetic dimensions represented by the four higher-order factors, a single genetic dimension should underlie each basic trait that is shared by other traits constituting the higher-order factor. Evidence of a genetic effect specific to each trait would be provided by evidence that the 18 basic traits are composed of two or more genetic dimensions.
A one-factor common pathways model did not provide a satisfactory fit for any of the 18 basic traits. On the other hand, an independent pathways model postulating a single genetic dimension explained the covariation among specific traits for 12 of the 18 basic trait scales: Anxiousness, Cognitive Dysregulation, Compulsivity, Conduct Problems, Identity Problems, Insecure Attachment, Intimacy Problems, Oppositionality, Rejection, Stimulus Seeking, Submissiveness, and Suspiciousness. The results of model fitting for illustrative scales are provided in Table 3.5. For three of these scales, Intimacy Problems, Rejection, and Stimulus Seeking, the common genetic dimension accounted for little of the variance for one or more of the specific trait scales, indicating that a specific genetic factor influenced these traits.Two genetic dimensions were found to underlie four scales: Affective Lability, Narcissism, Restricted Expression, and Social Avoidance. Three common genetic dimensions contributed to Callousness (see Table 3.5).
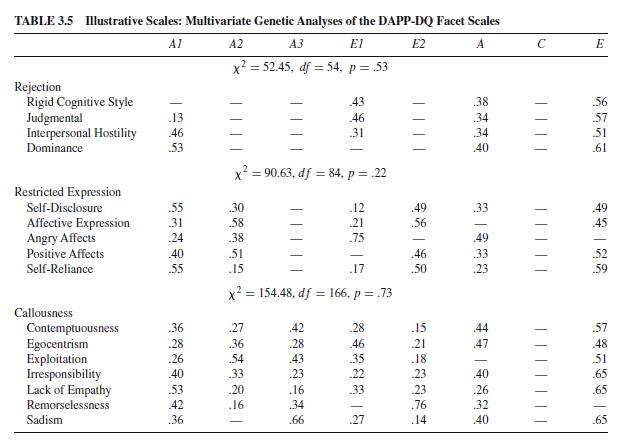
Multivariate analyses of normal and disordered personality traits suggest that multiple genetic and environmental factors influence the covariant structure of traits. They also confirm the findings of the regression analyses that many lower-order traits are influenced by one or more genetic dimensions specific to those traits. Finally, in both sets of analyses, the common pathways model did not provide a better fit to the data than did the independent pathways model. This suggests that the general genetic dimensions found by Livesley and colleagues (1998) and others by factor analyzing matrices of genetic correlations do not influence each trait through a latent phenotypic variable, but rather exert a direct influence on each trait.
Implications for Personality Structure
The studies described in the previous section reveal a complex genetic basis for personality. Multiple genetic dimensions differing in the breadth of their effects contribute to personality phenotypes (Jang et al., 1998; Livesley et al., 1998; Livesley & Jang, 2000). Some are relatively specific dimensions that influence single phenotypic traits, whereas others have broader effects influencing multiple phenotypically distinct but covarying traits. Consequently, many traits appear to be influenced by multiple genes and gene systems. Similarly, trait covariation seems to arise from multiple genetic effects. Genetic effects on traits appear to be direct rather than mediated by higher-order entities. These findings require replication. Nevertheless they appear to challenge models of personality positing links between specific genetically based neurotransmitter systems and specific personality traits. They also suggest a different conception of the trait hierarchies from that assumed by many trait taxonomies.
Hierarchical Structure
Factor analyses of genetic correlations and the modeling studies cited in the previous section identified general genetic factors that account for trait covariation. The model-fitting analyses also confirmed conclusions based on regression analyses that lower-order traits are not merely components of higher-order traits, but rather are distinct etiological entities. It appears that each basic or facet trait is influenced by general and specific genetic factors. Genetic dimensions that affect multiple traits appear to influence each trait directly rather than indirectly through a higher-order phenotypic entity. This raises questions about the basis for the hierarchy consistently identified by factor analytic studies and the conceptual status of higher-order constructs like neuroticism and extraversion and their role in theories of individual differences.
Although the facets delineating each of the five-factor domains covary due to shared genetic effects, it is not necessary to invoke a higher-order latent construct to explain this covariation. This raises the possibility that higher-order constructs such as neuroticism merely represent the pleiotropic action of genes. If this is the case, neuroticism and other higher-order domains are not entities that are distinct from the specific traits that delineate them. They are not traits in Allport’s sense of distinct phenotypic entities with an underlying biology, but rather heuristic devices that represent clusters of traits that covary because of a common genetic effect. This is consistent with the conception of domains as lexical categories (Saucier & Goldberg, 1996). Nevertheless, facet traits defining domains such as neuroticism and extraversion overlap sufficiently to justify grouping them into an overall global measure.
The model of trait structure implied by these findings differs from that of traditional trait theories. With traditional models in which lower-order traits are nested within a few higher-order factors, it follows that any statement about the higher-order factor applies to all subordinate traits. This is not the case with the model proposed because each basic trait has its own specific etiology. A second difference is that traditional hierarchical models seem to assume that trait taxonomies are similar to any classification based on set theory principles. At each level in the hierarchy, categories are assumed to be exhaustive and exclusive (Simpson, 1961). Exhaustiveness means that trait categories exist to classify all subordinate traits, whereas exclusiveness refers to the principle that each subordinate feature can be classified into only one superordinate trait. Considerable effort has been expended in attempts to delineate a structure with these properties. Indeed, this is the reason for debate on number and content of domains. It also explains Costa and McCrae’s insistence that domains are equal in breadth. If they are not, the five-factor model is open to the criticism that the model is not sufficiently parsimonious, as argued by Eysenck. This theoretical structure is understandable if trait taxonomies are conceptualized only as lexical structures. It is possible, however, that traits at the biological level are not organized in the systematic way proposed by the five-factor model.
There are no a priori reasons to assume that all basic traits must be organized into a hierarchy or that each higher-order domain is equally broad and defined by an equal number of facets as hypothesized by the five-factor model. An equally plausible model is that traits are organized into clusters that differ in the number of basic traits that they subsume and that the hierarchy is incomplete, with some specific traits showing minimal degrees of covariation. This structure is illustrated by the findings regarding the structure of the higher-order dimension of compulsivity identified in studies of personality disorder traits (Livesley et al., 1998). Pathways models identified a single genetic dimension underlying the specific traits that define this construct. Factor analyses show that it is consistently not related to other traits—hence, the three phenotypic traits that delineate compulsivity from separate higher-order factors. Compulsivity is, however, a trait narrower than other higher-order domains. It appears to represent a distinct basic or lower-order trait based on a single genetic dimension that does not have a hierarchical relationship with other basic traits.
Basic-Level Traits: Defining the Basic Unit of Personality
The idea that personality is inherited as a few genetic modules with broad effects and a large number of modules with more specific effects focuses attention on the significance of lower-order or basic traits. These findings are similar to evaluations of hierarchical models of cognitive ability that also provide evidence that specific abilities are heritable (Casto, DeFries, & Fulkner, 1995; Pedersen, Plomin, Nesselroade, & McClearn, 1992). Basic traits do not appear to be specific exemplars of the higher-order traits that they define or blends of two or more factors (Hofstee, DeRaad, & Goldberg, 1992). Rather, they are discrete genetic entities with their own biological basis. This suggests that personality models that reduce traits to a few global domains do not reflect the genetic architecture of normal or disordered personality. As noted earlier, personality research has tended to neglect these traits in favor of more global dimensions. Yet evidence of specificity of genetic effects suggests that the basic traits are the fundamental building blocks of personality that are more important for understanding personality than are the global constructs that have traditionally been the focus of research and explanation. This approach again raises the question of how basic traits should be conceptualized and defined, as well as which criteria are relevant to defining domains.
Costa and McCrae (1998) noted the challenges of delineating a comprehensive set of basic traits. The specificity of genetic effects also reveals the challenge involved because of the large number of genetic dimensions that are likely to be involved. A genetic perspective does, however, provide a definition of a basic dimension that could facilitate the identification and assessment of these traits. The usual psychometric criteria used to develop homogeneous scales could be supplemented with the genetic criterion that a basic trait scale represents a single specific genetic dimension. With this approach, items assessing a basic trait would form a genetically homogeneous unit as opposed to a factorially homogeneous unit. Items could then be selected according to their correlation with the underlying genetic dimension. Thus items forming a scale would share the same general and specific genetic etiology. With this approach, the goal would be to use behavioral genetic techniques to bring about definitions of the phenotype that correspond to what Farone, Tsuang, and Tsuang (1999) refer to as “genetically crisp categories” (p. 114).
An example of this approach is provided by a study of the genetic structure of the Eysenck Personality Questionnaire (Heath, Eaves, & Martin, 1989). This instrument has three broad scales composed of 21 to 25 items that assess Neuroticism, Extraversion, and Psychoticism. Heath and colleagues extracted a common genetic and environmental factor for Neuroticism and Extraversion, indicating that these items are etiologically homogeneous. In contrast, little evidence was found for a common genetic factor for the Psychoticism items. Subsequent analyses showed that the items formed into two distinct genetic factors: paranoid attitudes and hostile behavior. The results of such a systematic evaluation of item etiology could be used to form etiologically homogeneous scales.
This approach could be used either to develop new scales or modify existing scales so that they resemble the underlying genetic architecture more closely. This could be achieved by applying differential weights that index the influence of specific genetic and environmental influences on different traits. In this way, questions about the phenotypic structure of personality are addressed, and scales could be constructed so that they do not reflect competing genetic and environmental influences.
The estimation of genetic and environmental factor scores is a relatively new and active area of research. Sham et al. (2001) recently described a method that permits these genetic factor scores to be computed. Their method uses the following equation:
y = γ Σ−1 x (3.2)
where y = factor score for the common genetic factor, ã = the factor loadings of each variable on the genetic factor of interest (i.e., the column vector of estimated path coefficients that represent the correlations between the common genetic or environmental factor and the observed measures), Σ−1 = correlation matrix between all of the variables (i.e., the inverse of the correlation matrix of the observed measures), and x = each person’s score or response to each of the variables (i.e., column vector of observed values on the measures). Other methods are also available to compute genetic and environmental factor scores (Thomis et al., 2000).
Domain Content
As discussed earlier, the facet structure of several five-factor domains is still unclear. The same behavioral genetic approach used to define and measure basic trait scales could also be applied to the delineation of domain content. The unity of a domain is demonstrated by evidence that a single common genetic factor influences all the facets composing the domain. This approach could be used to clarify the location of impulsivity within the higher-order structure. The five-factor model locates impulsivity in Neuroticism, whereas Eysenck places it within Extraversion. As noted earlier, the bivariate correlations of this facet with other Neuroticism facets assessed with the NEO-PI-R are lower than correlations between other facets. Etiological data could be used to relocate impulsivity with other traits with which it shares a common etiology. Alternatively the item content could be changed based on genetic and environmental etiology so that correlations with the other Neuroticism facets are increased (of the loadings on the common factors are increased). In the case of the DAPP scales, impulsivity is part of the phenotypic trait of stimulus seeking along with sensation seeking and recklessness. Multivariate genetic analyses showed that a single common genetic factor underlies this dimension that is defined by sensation seeking and recklessness (see Table 3.5). Impulsivity has a low loading on the factor and a substantial specific heritable component. It appears that impulsivity as defined within the DAPPstructure is a specific heritable entity and not the result of interaction between extraversion and constraint or psychoticism as suggested by Depue and Collins (1999) or extraversion and psychoticism as suggested by Gray (1970, 1973, 1987; Pickering & Gray, 1999), although it is consistent with Gray’s argument that impulsivity is a fundamental dimension of temperament.
The findings of behavioral genetic studies of personality structure also have implications for attempts to identify the putative genes for personality. Most molecular genetic studies of personality use an analytic strategy that correlates a total personality trait score such as Neuroticism with variations in the candidate allele (Lesch et al., 1996). As the studies described show, the total scale score confounds multiple genetic and environmental effects and reduces the power to detect putative loci. The use of etiological factor scores that index the proportions of the personality phenotype directly attributable to specific genetic and environmental effects (Boomsma, 1996; Sham et al., 2001; Thomis et al., 2000) could reduce these confounds.
Universality of Trait Structure
Most models of personality traits including Eysenck’s threefactor model (Eysenck & Eysenck 1992), the five-factor model, and diagnostic categories of personality disorder proposed in the DSM-IV (American Psychiatric Association, 1994) assume that the taxonomies proposed reflect a universal structure. This assumption is also assumed to apply to the measures developed to assess these constructs. The only differences that these models of personality (and their measures) permit between cultures and other groups (e.g., gender) are quantitative in nature; they typically mean differences in trait levels or severity. If these assumptions are correct, we should find that the etiological architecture of personality is also invariant across cultures and other basic groupings. We discuss this idea with respect to cross-cultural comparisons and the effects of gender.
Cross-Cultural Comparisons
Multiple studies show that the observed factorial structure of scales such as the NEO-PI-R is stable across cultures. For example, McCrae and Costa (1997) reported that the five-factor structure is consistent across samples from the United States, Western Europe, and Asia (see also Costa & McCrae, 1992; McCrae et al., 2000). The issue of cross-cultural stability also applies to etiological structure. Earlier, we described fitting an independent pathways model to the six facets defining NEO-PI-R domains in independent samples of German and Canadian twins. The universality of genetic effects can be evaluated by testing the equivalence of the genetic and environmental structures across independent samples. It is possible to test whether: (a) the same genetic and environmental factors influenced the Canadian and German samples; and (b) whether these factors influenced each sample to the same degree. Two tests of equivalency were applied. The first evaluated equivalency of model form by testing the hypothesis that the same kind and number of genetic parameters are required to explain the data across the two samples. Sample differences are hypothesized to be limited to differences in the magnitude of the genetic and environmental influence exerted on a domain’s facet scales. If equivalence of model form was supported across the samples, the next step was to evaluate the magnitude of genetic and environmental influences across samples. This was accomplished by applying a model with the same parameters to both samples. That is, the model specified the same number and type of factors in both samples and identical and constrained the factor loadings to be identical.
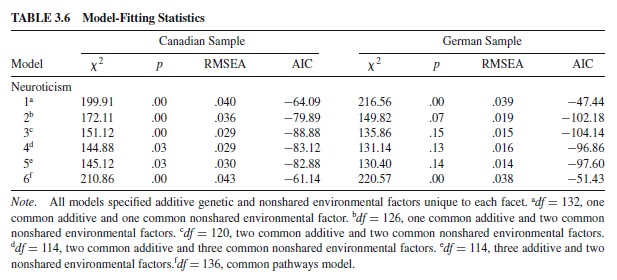
The results of tests of model form and magnitude for NEO-PI-R Neuroticism are shown in Table 3.6. The same number and types of genetic and environmental influences (two additive genetic and two nonshared environmental common factors) were identified in both samples, suggesting that the structure of neuroticism was similar across the samples. When the factor loadings on the common factors from the German sample were made to be the same as those on the Canadian sample (and vice versa), the model no longer fit the data. The results suggested that the primary differences between the German and Canadian samples were limited to the magnitude rather than kind of genetic and environmental effects supporting the claim that the factorial structure of the NEO-PI-R facets is universal.
Gender Differences
Personality tests are usually constructed to minimize genderbased differences by eliminating items whose intercorrelations with the other items can be attributable to gender and eliminating items evoking marked gender differences in endorsement. The approach yields scales that are applicable to both females and males but it overlooks the possibility of gender differences in the etiology. Behavioral genetic methods may be used to determine whether the same genetic and environmental factors influence personality measure scores in males and females and whether the etiological architecture underlying the factorial structure of a personality measure is the same in males and females.
The first question can be answered by fitting sex-limitation models to personality data (Neale & Cardon, 1992). This is accomplished by fitting a simple extension of the usual heritability model that uses data from same- and opposite-sex twin pairs to test whether the same genetic factors operate in males and females. In this case, gender differences are limited to differences in the magnitude of genetic and environmental influences. Another form of sex-limited gene expression occurs when different genes control the expression of a trait that is measured in the same way in males and females. With this form of sex-limitation, it is also possible to determine whether the same genes are present in both sexes but only expressed in one sex. This is evaluated by comparing the similarities of opposite-sex DZ twin pairs with same-sex DZ pairs. Sex-specific genetic influences are suggested when the similarity of opposite-sex pairs is significantly less than the similarities of male or female DZ pairs. The difference in the correlation is attributable to the gender composition of each zygosity group. When the same and opposite-sex DZ correlations are similar, gender differences are not indicated.
Only a few studies have investigated sex-limited gene expression in normal personality. The most notable is Finkel and McGue’s (1997) study that showed that the same genetic loci influence 11 out of the 14 scales of Multidimensional Personality Questionnaire (MPQ; Tellegen, 1982) in males and females. The heritable influences on the remaining three traits—Alienation, Control, and Absorption—indicated that the genetic influences were gender-specific. Jang, Livesley, and Vernon (1998) reported some evidence for sex-limited gene expression in 18 traits delineating personality disorder measured by the DAPP. All dimensions except Submissiveness in males, and Cognitive Dysfunction, Compulsivity, Conduct Problems, Suspiciousness, and Self-Harm in females were significantly heritable. Sex-by-genotype analyses suggested that the genetic influences underlying all but four DAPP dimensions (Stimulus Seeking, Callousness, Rejection, Insecure Attachment) were specific to each gender, whereas environmental influences were the same in both genders across all dimensions. Furthermore, the four higher-order dimensions derived from the 18 basic traits (Livesley et al., 1998) were also heritable across sex, and genetic effects were in common to both genders; the exception was Dissocial Behavior, which was not heritable in females.
Such evidence of sex-limited effects challenges the assumed universality of trait taxonomies. However, it could be argued that the results based on the DAPP and MPQ are atypical. The DAPP is a specialized scale designed primarily to assess personality dysfunction. The scale does not cover such areas of normal personality as Openness to Experience (Jang & Livesley, 1999; Schroeder et al., 1992) because abnormal variants of Openness are not included in clinical descriptions of personality disorder. The MPQ, unlike other scales, routinely reveals nonadditive genetic effects due to dominance (Waller & Shaver, 1994). This suggests that it may assess content different from that tested by scales such as the NEO-PI-R, which reveals genetic effects that are additive (e.g., Jang et al., 1998).
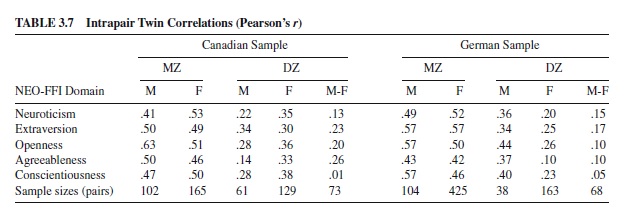
A more appropriate evaluation of the assumption of universality would be to examine sex-limited gene expression on a major model of personality such as the five-factor model. Evaluation of whether the same genes are present across different samples is similar to the evaluation of crosscultural effects. Jang, Livesley, Riemann, and Angleitner (in press) applied sex-limitation models to NEO-FFI data obtained from the Canada and German twin samples described earlier. Two general models were fit to the data. The first specified additive genetic and nonshared environmental influences for females and males and a male-specific genetic factor. The second tested whether heritable influences common to males and females were the same across the two samples. Table 3.7 reports the intrapair twin correlations for each zygosity group in each sample. The MZ male and MZ female correlations exceed their respective DZ correlations, suggesting the presence of heritable influences on each NEO-FFI domain in each sample. Of particular interest is the comparison between the DZ opposite-sex correlations and the same-sex DZ correlations. In both samples, the DZ oppositesex correlation for Conscientiousness was near zero, suggesting the presence of differential gender effects. The final form of the best-fitting model is presented in Table 3.8. The results suggest that genetic and environmental influences common to males and females influence four of the five FFM domains. The exception was Conscientiousness, for which genderspecific additive genetic influences operate. However, the external events and experiences specific to each twin— nonshared environmental influences—are common to males and females. The results also suggest that the type and magnitude of genetic and environmental influence were the same across the two groups, supporting the notion that the fivefactor model as assessed by the NEO-FFI is applicable to different cultures and genders.

This study has several limitations. The first is that the sample sizes are rather small in both samples, especially male DZ twin pairs and opposite-sex pairs. The twin covariances associated with these two zygosity types, especially the oppositesex pairs, are crucial for the validity of the analyses. The availability of relatively few twin pairs calls into question the stability of the correlations and thus the detection of sexlimited genes—as was obtained for Conscientiousness. Second, the study used the NEO-FFI, the short form of the NEO-PI-R. The full scale might produce different results because long versions of these scales sample domains more thoroughly. As such, the present results should be considered tentative until replicated on a larger sample using full-scale versions, as well as other measures of personality.
These analyses suggest that although most personality traits are influenced by the same genes in both genders (the implication being, e.g., DRD4 influences novelty seeking in both men and women), this is not true for all traits. The previous section suggested several explanations, but it is also possible that at the molecular level, different genes (or yet-tobe-discovered polymorphisms) differentially influence personality across genders. If this is the case, the genetic and environmental architecture of some scales may differ by gender. This could be evaluated by fitting independent and common pathways models to data from sister pairs and brother pairs separately and constraining the models (in form and magnitude) to be equal across gender groups. The sex-limitation model described previously that uses data from brothersister pairs to test for gender-specific effects can be expanded to the multivariate case to further explore gender differences in personality. As far as we are aware, few multivariate genetic analyses of gender differences have been conducted, probably because many studies have limited data collection to sister pairs or have difficulty obtaining data from brother pairs (Lykken, McGue, & Tellegen, 1987).
Environmental Effects
Although our primary concern is with the genetic basis for personality structure, any discussion of genetic influence would be incomplete without reference to environmental factors. Twin studies consistently show that about 50% of the variance in personality traits is explained by environmental factors and that most of this is accounted for by nonspecific influences; common environmental influences do not appear to contribute to personality variation (Plomin & Daniels, 1987). This etiological model derived from twin studies is confirmed by a large-scale study of Neuroticism by Lake, Eaves, Maes, Heath, and Martin (2000) that showed that individual differences in neuroticism were not transmitted from parent to offspring via the environment but rather by genetic factors. The size and unique features of their data set (45,880 twin pairs and their relatives on two continents) allowed them to test models of genetic transmission as well as geneenvironment correlations. The results suggest that the environment exerts a contemporaneous influence on individual differences in neuroticism. That is, its effects are located in the current environment as opposed to being preset like genetic factors that are passed to individuals from their parents.
Although nonshared environmental factors are important, the nature of these variables and the way they affect personality remain unclear. Despite considerable research effort (e.g., Hetherington, Reiss, & Plomin, 1994; Turkheimer & Waldron, 2000) using a variety of methods (Baker & Daniels, 1990; Hetherington et al., 1994; Reiss et al., 1994; Vernon, Lee, Harris, & Jang, 1996) the results have been uniformly disappointing: Few nonshared influences on personality have been identified (Turkheimer & Waldron, 2000). Most studies have, however, investigated the effects of the nonshared environment on the single variables; few studies have examined the effects of the nonshared environment on trait covariance. The study by McCrae et al. (in press) and the illustrative multivariate genetic analyses of the NEO-PI-R and DAPP presented earlier suggest that the nonshared environmental factors have an influence on personality structure different from that of genetic factors. They do not appear to contribute to trait substantially to the trait covariation described by trait taxonomies.
It also appears that the environment does not have an effect that is independent of preexisting genetic factors. Reviews by Reiss, Neiderhiser, Hetherington, and Plomin (2000) and Caspi and Bem (1990) document personalityenvironment interaction and the way the individuals select and create their own environment. Genetic factors influence the environmental variables that are the focus of attention and the situations that the individual selects. For example, some kinds of life events are not independent of the individual; rather, their occurrence is influenced by such traits as Neuroticism and Extraversion (e.g., Poulton & Andrews, 1992; Magnus, Diener, Fujita, & Pavot, 1993). Saudino, Pedersen, Lichtenstein, and McClearn (1997) showed that all genetic variance on controllable, desirable, and undesirable life events in women was common to the genetic influences underlying EPQ Neuroticism and Extraversion, NEO-FFI Openness to Experience (Costa & McCrae, 1985). Genetic influences underlying personality scales had little influence on uncontrollable life events because this variable was not heritable. Kendler and Karkowski-Shuman (1997) showed that the genetic risk factors for major depression increased the probability of experiencing significant life events in the interpersonal and occupational-financial domains, probably because individuals play an active role in creating their own environments. Heritable factors, such as personality and depression, influence the types of environments sought or encountered. Jang, Vernon, and Livesley (2000) report significant genetic correlations between the Family Environment Scale (FES: Moos & Moos, 1974) subscale of Cohesiveness and DAPP higher-order factors of Emotional Dysregulation (−.45), Inhibition (−.39), FES Achievement Orientation and DAPP Dissocial Behavior (.38), Inhibition (−.58), and FES Intellectual Cultural Orientation and DAPP Emotional Dysregulation (−.34). These results help to explain why measures of the environment often have a heritable component: They often reflect genetically influenced traits (Saudino et al., 1997).
Using the factor score approach described earlier, Thomis et al. (2000) computed genetic factor scores for measures of muscle strength obtained from a sample of MZ and DZ twins. The twins were then subjected to a 10-week muscle strength training regimen. The muscle strength genetic factor scores explained the greatest proportion of the variance pre- and posttraining, indicating that genes are switched on, so to speak, in response to stress due to training, thus demonstrating the existence of gene-environment interaction. Findings such as these suggest that the environmental factors that influence traits are partially dependent on preexisting genetically based personality traits. For example, a person scoring highly on a genetically based trait like sensation seeking will seek out environments conducive to the expression of this personality genotype, such as engaging exciting sports. For this reason, molecular genetic studies designed to identify the genes for personality need to incorporate measures of personality that separate the effects of genes and environment on the phenotype.
Molecular Genetics
From a genetic perspective, dimensions of individual differences in personality are complex traits.That is, multiple genes and gene systems and multiple environmental factors influence each trait (Plomin, DeFries, McClearn, & McGuffin, 2000). The emergence of molecular genetics prompted considerable optimism about the possibility of identifying the genetic component or quantitative trait loci (QTLs) of these traits. Such a development would radically change the nature of personality research by enabling investigators to link behavioral dimensions to underlying molecular genetic structures. This would provide a more powerful way to resolve trait taxonomic issues that the behavioral genetic approaches discussed. The results of such studies have, however, been inconsistent, replications have often failed, and progress has been slower than expected.
One of the earliest studies investigated the relationship between Novelty Seeking and dopamine D4 or DRD4 receptor (Cloninger, Adolfsson, & Svrakic, 1996). Earlier Cloninger (1987; Cloninger, Svrakic, & Przybeck, 1993) proposed a model of personality that postulated that the expression of each personality trait is modulated by a specific genetically controlled neurotransmitter system. Specifically, Novelty Seeking is controlled by the dopaminergic system, Harm Avoidance by the serotonin system, and Reward Dependence by norephinephrine. Cloninger and colleagues (1996) reported a polymorphism of the D4 receptor that accounted for about 10% of the variance. Several replications have been reported (Benjamin, Greenberg, & Murphy, 1996; Ebstein, Novick, & Umansky, 1996; Ebstein, Segman, & Benjamin, 1997) along with many failed replications (Ebstein, Gritsenko, & Nemanov, 1997; Malhotra, Goldman, Ozaki, & Breier, 1996; Ono et al., 1997; Pogue-Geile, Ferrell, Deka, Debski, & Manuck, 1998; Vandenbergh, Zonderman, Wang, Uhl, & Costa, 1997).
Similarly, several studies have demonstrated a relationship between the serotonergic system and Harm Avoidance, Neuroticism, or related constructs (Hansenne & Ansseau, 1999; Rinne, Westenberg, den Boer, & van den Brink, 2000), and significant associations were reported with the serotonin transporter gene, 5-HTTLPR (Katsuragi et al., 1999). However, several studies have failed to replicate these findings (Flory et al., 1999; Gelernter, Kranzler, Coccaro, Siever, & New, 1998; Hamer et al., 1999; Herbst, Zonderman, McCrae, & Costa, 2000). Gustavsson et al. (1999) also failed to replicate these findings using the Karolinska Scales of Personality.
These inconsistencies can be attributed to conceptual and measurement issues. The early studies in particular were often based on a conceptual model that assumed that personality is influenced by relatively few genes, each accounting for substantial variance. As noted, the evidence does not support this approach. There has also been a tendency to assume that each trait was linked a specific neurotransmitter system. More recently, however, attention has focused on pleiotropic effects by investigating the possibility that a given polymorphism influences several traits. Work on the serotonin transporter gene, for example, suggests that it is not associated with a single trait but rather has a pleiotropic relationship with Neuroticism and Agreeableness. Studies on humans and primates suggest that altered serotonin activity is related to negative emotional states such as depression, anxiety, and hostility, and to social behaviors such as dominance, aggression, and affiliation with peers (Graeff, Guimaraes, De Andrade, & Deakin, 1996; Knutson et al., 1998; Murphy et al., 1998). Knutson and colleagues (1998) found that administration of the specific serotonin reuptake inhibitor, paroxetine, decreased negative affect and increased social affiliation in normal human subjects. Lesch and colleagues (1996) reported that individuals carrying the 5-HTTLPR-S allele had increased total scores on NEO-PI-R Neuroticism and the facets of Anxiety, Angry Hostility, Depression, and Impulsiveness. The allele accounted for 3 to 4% of the total variance in these scales. Unexpectedly, the allele was also associated with a decreased NEO-PI-R Agreeableness score. Greenberg et al. (1999) recently replicated these findings. Hamer et al. (1999) showed that 5-HTTLPR-S genotypes were significantly associated with increased Harm Avoidance (which correlates .66 with NEO-PI-R Neuroticism) and decreased Self-Directedness (correlated −.64 with NEO-PI-R Neuroticism), Reward Dependence, and Cooperativeness (shown to correlate .43 and .66 with NEO-PI-R Agreeableness). These effects accounted for .80%, 1.98%, .97%, and 2.60% of the total variance in these scores, respectively. Mazzanti et al. (1998), Peirson et al. (2000), and Benjamin et al. (2000) have reported replications.
Measurement problems contributing to inconsistent findings include the use of measures with less-than-optimal psychometric properties and the use of relatively broad personality constructs. Comparison of the dopamine–novelty seeking and serotonin-neuroticism studies suggests that the serotonin-neuroticism literature is less ambiguous than the dopamine–novelty seeking literature. These differences appear to be related to scale properties. Inconsistent findings may also be due to the confounding of genetic and environmental influences on the phenotypes. As we have tried to show, many constructs and scales are etiologically heterogeneous.
Twin studies estimating statistical pleiotropy could contribute to molecular genetic studies by identifying traits that are etiologically homogeneous units and etiologically related. Molecular genetic work could then be used to confirm these associations by identifying the actual genes that account for trait covariance. This would provide the strongest basis for revising personality models and allocating traits to etiologically related domains.
Conclusions
The thesis of this research paper is that behavioral genetic approaches promise to provide an additional perspective that may help to resolve some of the more intractable problems in delineating and conceptualizing personality structure. The evidence reviewed suggests an alternative perspective on the trait structure of personality that complements traditional conceptions. Although trait theory has largely concentrated on mapping personality in terms of broad global traits, the evidence suggests that personality is inherited as a large number of genetic dimensions that have relatively specific effects on personality phenotypes and a smaller number of genetic dimensions that have broader effects, perhaps through a modulating influence on related dispositions. These dimensions with broader effects appear to account for some of the observed covariation among traits. They do not appear, however, to exert these effects through higher-order phenotypic structures, but rather through a direct influence on each basic trait. We assume that these common features are more likely to involve modulating functions or common mechanisms that regulate each trait in a given cluster.
These tentative conclusions suggest the need to reconsider traditional models of the hierarchical structure of personality in which traits are organized into broad domains due to the effects on broad dispositions. Instead, the organization of traits into clusters is assumed to arise from the pleiotropic effects of genetic dimensions that affect multiple traits. Under these circumstances, it is conceivable that not all traits are organized into clusters of covarying features, but rather remain relatively distinct characteristics. Nor is it inevitable the traits are hierarchically organized in similar ways across domains. That is, it is possible that the symmetrical hierarchical structure avidly sought by trait theorists and students of psychopathology does not reflect the way personality is organized at a genetic level. An equally feasible structure would involve considerable differences in complexity across domains. Some domains may consist of a relatively large number of traits, whereas others may consist of only one or two genetically homogeneous traits. These assumptions are consistent with the lexical view of Saucier and Goldberg (1996), who argued that the five domains are merely a convenient way of organizing lower-order traits and that there is no inherent reason to assume that domains are equal in breadth or in pervasiveness.
Although behavioral genetic analyses show that environmental factors exert a considerable influence on personality, they do not appear to influence the structural relationships among traits to any appreciable extent. Instead, environmental factors appear to exert a more contemporaneous effect on trait expression. The nature of these factors and the way that they function remain important topics of research.
Bibliography:
- Allport, G. W. (1961). Pattern and growth in personality. New York: Holt, Reinhart and Winston.
- Almagor, M., Tellegen, A., & Waller, N. G. (1995). The big seven model: Across-cultural replication and further exploration of the basic dimensions of natural language trait descriptors. Journal of Personality and Social Psychology, 69, 300–307.
- American Psychiatric Association (1994). The diagnostic and statistical manual for mental disorders (4th ed.). Washington, DC: Author.
- Ashton, M. C., Lee, K., Vernon, P. A., & Jang, K. L. (2000). Fluid intelligence, crystallized intelligence, and the Openness/Intellect factor. Journal of Research in Personality, 34, 198–207.
- Avia, M. D., Sanz, J., Sanchez-Bernardos, M. L., Martinez-Arias, M. R. Silva, & Graña, (1995). The five-factor model—II. Relations of the NEO-PI with other personality variables. Personality and Individual Differences, 19, 81–97.
- Baker, L. A., & Daniels, D. (1990). Nonshared environmental influences and personality differences in adult twins. Journal of Personality and Social Psychology, 58, 103–110.
- Benjamin, J., Greenberg, B., & Murphy, D. L. (1996). Mapping personality traits related to genes: Population and family association between the D4 dopamine receptor and measures of novelty seeking. Nature Genetics, 12, 81–84.
- Benjamin, J., Osher, Y., Lichtenberg, P., Bachner-Melman, R., Gritsenko, I., Kotler, M., Belmaker, R. H., Valsky, V., Drendel, M., & Ebstein, R. P. (2000). An interaction between the catechol O-methyltransferase and serotonin transporter promoter region polymorphisms contributes to Tridimensional Personality Questionnaire persistence scores in normal subjects. Neuropsychobiology, 41, 48–53.
- Bergeman, C. S., Chipeur, H. M., Plomin, R., Pederson, N. L., McClearn,G.E.,Nesselroade,J.R.,Costa,P.T.,&McCrae,R.R. (1993). Genetic and environmental effects on openness to experience, agreeableness, and conscientiousness: An adoption/twin study. Journal of Personality, 61, 159–179.
- Boomsma, D. I. (1996). Using multivariate genetic modeling to detect pleiotropic quantitative trait loci. Behavior Genetics, 26, 161–166.
- Bouchard, T. J. (1999). Genes, environment, and personality. In S. J. Ceci & W. M. Williams (Eds.), The nature-nurture debate: The essential readings. Essential readings in developmental psychology (pp. 97–103). Malden, MA: Blackwell.
- Carey, G. (1987). Is extraversion a unitary trait? Behavior Genetics, 17,
- Carey, G., & DiLalla, D. L. (1994). Personality and psychopathology: Genetic perspectives. Journal of Abnormal Psychology, 103, 32–43.
- Caspi, A., & Bem, D. J. (1990). Personality continuity and change across the life course. In L. A. Pervin (Ed.), Handbook of personality: Theory and research. New York: Guilford.
- Casto, S. D., DeFries, J. C., & Fulker, D. W. (1995). Multivariate genetic analysis of Wechsler Intelligence Scale for Children— Revised (WISC–R) factors. Behavior Genetics, 25, 25–32.
- Cloninger, C. R. (1987). A systematic method for clinical description and classification of personality variants: A proposal. Archives of General Psychiatry, 44, 573–588.
- Cloninger, C. R., Adolfsson, R., & Svrakic, N. M. (1996). Mapping genes for human personality. Nature Genetics, 12, 3–4.
- Cloninger, C. R., Svrakic, D. M., & Przybeck, T. R. (1993). A psychobiological model of temperament and character. Archives of General Psychiatry, 50, 975–990.
- Costa, P. T., & McCrae, R. R. (1985). Manual for the NEO Personality Inventory. Odessa, FL: Psychological Assessment Resources.
- Costa, P. T., & McCrae, R. R. (1992). Revised NEO Personality Inventory (NEO-PI-R) and the NEO Five-Factor Inventory (NEO-FFI) professional manual. Odessa, FL: Psychological Assessment Resources.
- Costa, P. T., & McCrae, R. R. (1995). Domains and facets: Hierarchical personality assessment using the Revised NEO Personality Inventory. Journal of Personality Assessment, 64, 21–50.
- Costa, P. T., & McCrae, R. R. (1997). Longitudinal stability of adult personality. In R. Hogan & J. A. Johnson (Eds.), Handbook of personality psychology (pp. 269–290). San Diego, CA: Academic Press.
- Costa, P. T., & McCrae, R. R. (1998). Six approaches to the explication of facet-level traits: Examples from conscientiousness. European Journal of Psychology, 12, 117–134.
- Crawford, C. B., & DeFries, J. C. (1978). Factor analysis of genetic and environmental correlation matrices. Multivariate Behavioral Research, 13, 297–318.
- DeFries, J. C., & Fulker, D. W. (1986). Multivariate behavioral genetics and development: An overview. Behavior Genetics, 16, 1–10.
- Depue, R. A., & Collins, P. F. (1999). Neurobiology of the structure of personality: Dopamine, facilitation of incentive motivation, and extraversion. Behavioral and Brain Sciences, 22, 491– 569.
- Digman, J. M. (1990). Personality structure: Emergence of the fivefactor structure. Annual Review of Psychology, 41, 417–440.
- Digman, J. M., & Inouye, J. (1986). Further specification of the five robust factors of personality. Journal of Personality and Social Psychology, 50, 116–123.
- Draycott, S. G., & Kline, P. (1995). The Big Three or the Big Five—the EPQ-R vs. the NEO-PI: A research note, replication and elaboration. Personality and Individual Differences, 18, 801–804.
- Ebstein, R. P., Gritsenko, I., & Nemanov, L. (1997). No association between the serotonin transporter gene regulatory region polymorphism and the tridimensional personality questionnaire (TPQ) temperament of harm avoidance. Molecular Psychiatry, 2, 224–226.
- Ebstein, R. P., Novick, O., & Umansky, R. (1996). D4DR exon III polymorphism associated with the personality trait of novelty seeking in normal human volunteers. Nature Genetics, 12, 78–80.
- Ebstein, R. P., Segman, R., & Benjamin, J. (1997). 5–HT2C (HTR2C) serotonin receptor gene polymorphism associated with the human personality trait of reward dependence: Interaction with dopamine D4 receptor (D4DR) and dopamine D3 (D3DR) polymorphisms. American Journal of Medical Genetics, 74, 65–72.
- Eysenck, H. J. (1991). Dimensions of personality: 16, 5 or 3? Criteria for a taxonomic paradigm. Personality and Individual Differences, 12, 773–790.
- Eysenck, H. J. (1992). Areply to Costa and McCrae: P or Aand C— the role of theory. Personality and Individual Differences, 13, 867–868.
- Eysenck, H. J., & Eysenck, S. B. G. (1992). Manual for the Eysenck Personality Questionnaire–Revised. San Diego, CA: Educational and Industrial Testing Service.
- Farone, S. V., Tsuang, M. T., & Tsuang, D. W. (1999). Genetics of mental disorders. New York: Guilford Press.
- Finkel, D., & McGue, M. (1997). Sex differences and nonadditivity in heritability of the Multidimensional Personality Questionnaire Scales. Journal of Personality and Social Psychology, 72, 929–938.
- Flory, J .D., Manuck, S. B., Ferrell, R. E., Dent, K. M., Peters, D. G., & Muldoon, M. F. (1999). Neuroticism is not associated with the serotonin transporter (5–HTTLR) polymorphism. Molecular Psychiatry, 4, 93–96.
- Gelernter, J., Kranzler, H., Coccaro, E. F., Siever, L. J., & New, A. S. (1998). Serotonin transporter protein gene polymorphism and personality measures in African American and European American subjects. American Journal of Psychiatry, 155, 1332–1338.
- Goldberg, L. R., (1990). An alternative “description of personality”: The Big-Five factor structure. Journal of Personality and Social Psychology, 59, 1216–1229.
- Goldberg, L. R. (1993). The structure of phenotypic personality traits. American Psychologist, 48, 26–34.
- Gough, H. G. (1989). The California Psychological Inventory. In C. S. Newmark (Ed.), Major psychological assessment instruments (Vol. 2, pp. 67–98). Needham Heights, MA: Allyn & Bacon.
- Graeff, F. G., Guimaraes, F. S., De Andrade, T. G., & Deakin, J. F. (1996). Role of 5–HT in stress, anxiety, and depression. Pharmacology, Biochemistry, and Behavior, 54, 129–141.
- Gray, J. A. (1970). The psychophysiological basis of intraversionextraversion. Behavior Research and Therapy, 8, 249–266.
- Gray, J. A. (1973). Causal theories of personality and how to test them. In J. R. Royce (Ed.), Multivariate analysis and psychological theory, (pp. 409–463). London: Academic Press.
- Gray, J. A. (1987). The neuropsychology of emotion and personality. In S. M. Stahl & S. D. Iversen (Eds.), Cognitive neurochemistry (pp. 171–190). Oxford, England: Oxford University Press.
- Greenberg, B. D., Tolliver, T. J., Huang, S. J., Li, Q., Bengel, D., & Murphy, D. L. (1999). Genetic variation in the serotonin transporter promoter region affects serotonin uptake in human blood platelets. American Journal of Medical Genetics, 88, 83–87.
- Gustavsson, J. P., Nöthen, M. M., Jonsson, E. G., Neidt, H., Forslund, K., Rylander, G., Mattila-Evenden, M., Sedvall, G. C., Propping, P., & Åsberg, M. (1999). No association between serotonin transporter gene polymorphisms and personality traits. American Journal of Medical Genetics, 88, 430–436.
- Hamer, D. H., Greenberg, B. D., Sabol, S. Z., & Murphy, D. L. (1999). Role of serotonin transporter gene in temperament and character. Journal of Personality Disorders, 13, 312–328.
- Hampson, S. E., John, O. P., & Goldberg, L. R. (1986). Category breadth and hierarchical structure in personality: Studies of asymmetries in judgments of trait implications. Journal of Personality and Social Psychology, 51, 37–54.
- Hansenne, M., & Ansseau, M. (1999). Harm avoidance and serotonin. Biological Psychology, 51, 77–81.
- Heath, A. C., Eaves, L. J., & Martin, N. G. (1989). The genetic structure of personality: III. Multivariate genetic item analysis of the EPQ scales. Personality and Individual Differences, 10, 877–888.
- Heath, A. C., Neale, M. C., Kessler, R. C., Eaves, L. J., & Kendler, K. S. (1992). Evidence for genetic influences on personality from self-reports and informant ratings. Journal of Personality and Social Psychology, 63, 85–96.
- Herbst, J. H., Zonderman, A. B., McCrae, R. R., & Costa, P. T. (2000). Do the dimensions of the Temperament and Character Inventory map a simple genetic architecture? Evidence from molecular genetics and factor analysis. American Journal of Psychiatry, 157, 1285–1290.
- Hetherington, E. M., Reiss, D., & Plomin, R. (Eds.) (1994). Separate social worlds of siblings: The impact of nonshared environment on development. Hillsdale, NJ: Lawrence Erlbaum.
- Hofstee, W. K., deRaad, B., & Goldberg, L. R. (1992). Integration of the Big Five and circumplex approaches to trait structure. Journal of Personality and Social Psychology, 63, 146–163.
- Jackson, D. N. (1984). The Personality Research Form. Ontario, Canada: Sigma Assessment Systems.
- Jang, K. L., & Livesley, W. J. (1999).Why do measures of normal and disordered personality correlate? A study of genetic comorbidity. Journal of Personality Disorders, 13, 10–17.
- Jang, K. L., Livesley, W. J., Riemann, R., &Angleitner,A. (in press). The Five-Factor Model of personality: A behavioural genetic analysis of gender across cultures. Personality and Individual Differences.
- Jang, K. L., Livesley, W. J., Vernon, P. A., & Jackson, D. N. (1996). Heritability ofpersonality disorder traits: A twin study. Acta Psychiatrica Scandinavica, 94, 438–444.
- Jang, K. L., McCrae, R. R., Angleitner, A., Riemann, R., & Livesley, W. J. (1998). Heritability of facet-level traits in a cross-cultural twin study: Support for a hierarchical model of personality. Journal of Personality and Social Psychology, 74, 1556–1565.
- Jang, K. L., Vernon, P. A., & Livesley, W. J. (2000). Personality disorder traits, family environment, and alcohol misuse: A multivariate behavioural genetic analysis. Addiction, 95, 873– 888.
- John, O. P., & Srivastava, S. (1999). The big five trait taxonomy: History, measurement, and theoretical perspectives. In L. A. Pervin & O. P. John (Eds.), Handbook of Personality (pp. 102–138). New York: Guilford.
- Katsuragi, S., Kunugi, A. S., Sano, A., Tsutsumi, T., Isogawa, K., Nanko, S., & Akiyoshi, J. (1999). Association between serotonin transporter gene polymorphism and anxiety-related traits. Biological Psychiatry, 45, 368–370.
- Kendler, K. S., & Karkowski-Shuman, L. (1997). Stressful life events and genetic liability to major depression: Genetic control of exposure to the environment? Psychological Medicine, 27, 539–547.
- Knutson, B., Wolkowitz, O. M., Cole, S. W., Chan, T., Moore, E. A., Johnson, R. C., Terpstra, J., Turner, R. A., & Reus, V. I. (1998). Selective alteration of personality and social behavior by serotonergic intervention. American Journal of Psychiatry, 155, 373–379.
- Lake, R. I. E., Eaves, L. J., Maes, H. H., Heath, A. C., & Martin, N. G. (2000). Further evidence against the environmental transmission of individual differences in neuroticism from a collaborative study of 45,850 twins and relatives on two continents. Behavior Genetics, 30, 223–233.
- Lesch, K. P., Bengel, D., Heils, A., Zhang Sabol, S., Greenberg, D., Petri, S., Benjamin, J., Muller, C. R., Hamer, D. H., & Murphy, D. L. (1996). Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science, 274, 1527–1530.
- Livesley, W. J. (1986). Trait and behavioral prototypes of personality disorder. American Journal of Psychiatry, 143, 728–732.
- Livesley, W. J., & Jackson, D. N. (in press). Manual for the dimensional assessment of personality pathology. Port Huron, MI: Sigma Press.
- Livesley, W. J., Jackson, D. N., & Schroeder, M. L. (1992). Factorial structure of traits delineating personality disorders in clinical and general population samples. Journal of Abnormal Psychology, 101, 432–440.
- Livesley, W. J., & Jang, K. L. (2000). Toward an empirically based classification of personality disorder. Journal of Personality Disorders, 14, 137–151.
- Livesley, W. J., Jang, K. L., & Vernon, P. A. (1998). The phenotypic and genetic architecture of traits delineating personality disorder. Archives of General Psychiatry, 55, 941–948.
- Loehlin, J. C. (1987). Heredity, environment, and the structure of the California Psychological Inventory. Multivariate Behavioral Research, 22, 137–148.
- Loehlin, J. C. (1992). Genes and environment in personality development. Newbury Park, CA: Sage.
- Loehlin, J. C., & Nicholls, R. C. (1976). Heredity, environment, and personality. Austin: University of Texas Press.
- Lykken, D. T., McGue, M., & Tellegen, A. (1987). Recruitment bias in twin research: The rule of two-thirds reconsidered. Behavior Genetics, 17, 343–362.
- Magnus, K., Diener, E., Fujita, F., & Pavot, W. (1993). Extraversion and neuroticism as predictors of objective life events: A longitudinal analysis. Journal of Personality and Social Psychology, 65, 1046–1053.
- Malhotra, A. K., Goldman, D., Ozaki, N., & Breier, A. (1996). Lack of association between polymorphisms in the 5–HT-sub(2A) receptor gene and the antipsychotic response to clozapine. American Journal of Psychiatry, 153, 1092–1094.
- Mazzanti, C. M., Lappalainen, J., Long, J. C., Bengel, D., Naukkarinen, H., Eggert, M., Virkkunen, M., Linnoila, M., Goldman, D. (1998). Role of the serotonin transporter promoter polymorphism in anxiety-related traits. Archives of General Psychiatry, 55, 936–940.
- McArdle, J. J., & Goldsmith, H. H. (1990). Alternative common factor models for multivariate biometric analyses. Behaviour Genetics, 20, 569–608.
- McCrae, R. R., & Costa, P. T. (1985a). Comparison of EPI and psychoticism scales with measures of the five-factor model of personality. Personality and Individual Differences, 6, 587– 597.
- McCrae, R. R., & Costa, P. T. (1985b). Updating Norman’s “adequacy taxonomy”: Intelligence and personality dimensions in natural language and in questionnaires. Journal of Personality and Social Psychology, 49, 710–721.
- McCrae, R. R., & Costa, P. T. (1986). Clinical assessment can benefit from recent advances in personality psychology. American Psychologist, 41, 1001–1003.
- McCrae, R. R., & Costa, P. T. (1996). Toward a new generation of personality theories: Theoretical contexts for the five-factor model. In J. S. Wiggins (Ed.), The five factor model of personality (pp. 51–87). New York: Guilford.
- McCrae, R. R., Costa, P. T., & Piedmont, R. L. (1993). Folk concepts, natural language, and psychological constructs: The California Psychological Inventory and the five factor model. Journal of Personality, 61, 1–26.
- McCrae, R. R., & Costa, P. T., Jr. (1997). Personality trait structure as a human universal. American Psychologist, 52, 509–516.
- McCrae, R. R., Costa, P. T., Jr., Ostendorf, F., Angleitner, A., Hrebickova, M., Avia, M. D., Sanz, J., Sanchez-Bernardos, L., Kusdil, M. E., Woodfield, R., Saunders, P. R., & Smith, P. B. (2000). Nature over nurture: Temperament, personality, and life span development. Journal of Personality and Social Psychology, 78, 173–186.
- McCrae, R. R., Jang, K. L., Livesley, W. J., Riemann, R., & Angleitner, A. (in press). The five-factor model of personality: A behavioural genetic analysis of gender across cultures. Personality and Individual Differences.
- McCrae, R. R., & John, O. P. (1992). An introduction of the fivefactor model and its applications. Journal of Personality, 60, 175–215.
- Moos, R. H., & Moos, B. S. (1974). Manual for the Family Environment Scale. Palo Alto, CA: Consulting Psychologists Press.
- Murphy, D. L., Andrews, A. M., Wichems, C. H., Li, Q., Tohda, M., & Greenberg, B. (1998). Brain serotonin neurotransmission: An overview and update with an emphasis on serotonin subsystem heterogeneity, multiple receptors, interactions with other neurotransmitter systems, and consequent implications for understanding the actions of serotonergic drugs. Journal of Clinical Psychiatry, 59, 4–12.
- Neale, M. C., & Cardon, L. R. (1992). Methodology for genetic studies of twins and families. Dordrecht, Netherlands: Kulwer Academic Press.
- Ono, Y., Manki, H., Yomishura, K., Muramatsu, T., Higuchi, S., Yagi, G., Kanba, S., & Asai, M. (1997). Association between dopamine D4 receptor (D4DR) exon III polymorphism and novelty seeking in Japanese subjects. American Journal of Genetics, 47, 501–503.
- Passini, F. T., & Norman, W. T. (1966). A universal conception of personality structure? Journal of Personality and Social Psychology, 4, 44–49.
- Paunonen, S. V., & Jackson, D. N. (1996). The Jackson Personality Inventory and the Five factor model of personality. Journal of Research in Personality, 30, 42–59.
- Pedersen, N. L., Plomin, R., Nesselroade, J. R., & McClearn, G. E. (1992). A quantitative genetic analysis of cognitive abilities during the second half of the life span. Psychological Science, 3, 346–353.
- Peirson, A. R., Heuchert, J. W., Thomala, L., Berk, M., Plein, H., & Cloninger, C. R. (2000). Relationship between serotonin and the Temperament and Character Inventory. Psychiatry Research, 89, 29–37.
- Pickering, A. D., & Gray, J. A. (1999). The neuroscience of personality. In L. A. Pervin & O. P. John (Eds.), The handbook of personality (pp. 277–299). New York: Guilford.
- Plomin, R., & Caspi, A. (1998). DNA and personality. European Journal of Personality, 12, 387–407.
- Plomin, R., Chipeur, H. M., & Loehlin, J. C. (1990). Behavior genetics and personality. In L. A. Pervin (Ed.), Handbook of personality: Theory and research. New York: Guilford.
- Plomin, R., & Daniels, D. (1987). Children in the same family are very different, but why? Behavioral and Brain Sciences, 10, 44–59.
- Plomin, R., DeFries, J. C., & McClearn, G. E. (1990). Behavioral genetics: A primer (2nd ed.). New York: W. H. Freeman.
- Plomin, R., DeFries, J. C., McClearn, G. E., & McGuffin, P. (Eds.). (2000). Behavioral genetics: A primer (3rd ed.). New York: W. H. Freeman.
- Pogue-Geile, M., Ferrell, R., Deka, R., Debski, T., & Manuck, S. (1998). Human novelty seeking personality traits and dopamine D4 receptor polymorphisms: A twin and genetic association study. American Journal of Medical Genetics, 81, 44–48.
- Poulton, R. G., & Andrews, G. (1992). Personality as a cause of adverse life events. Acta Psychiatrica Scandinavica, 85, 35–38.
- Reiss, D., Neiderhiser, J. M., Hetherington, E. M., & Plomin, R. (2000). The relationship code: Deciphering genetic and social influences on adolescent development. Cambridge, MA: Harvard University Press.
- Riemann, R., Angleitner, A., & Strelau, J. (1997). Genetic and environmental influences on personality: A twin study. Journal of Personality, 65, 449–476.
- Rinne, , Westenberg, H. G. M., denBoer, J. A., & van den Brink, W. (2000). Serotonergic blunting to meta-chlorophenylpiperazine (m-CPP) highly correlates with sustained childhood abuse in impulsive and autoaggressive female borderline patients. Biological Psychiatry, 47, 548–556.
- Saucier, G., & Goldberg, L. R. (1996). The language of personality: Lexical perspectives on the five-factor model. In J. S.Wiggins (Ed.), The five factor model of personality (pp. 21–50). New York: Guilford.
- Saudino, K. J., Pedersen, N. L., Lichtenstein, P., & McClearn, G. E. (1997). Can personality explain genetic influences on life events? Journal of Personality and Social Psychology, 72, 196–206.
- Schroeder, M. L., Wormworth, J. A., & Livesley, W. J. (1992). Dimensions of personality disorder and their relationships to the big five dimensions of personality. Psychological Assessment: A Journal of Consulting and Clinical Psychology, 4, 47–53.
- Sham, P. C., Sterne, A., Purcell, S., Cherny, S., Webster, M., Rijsdijk, F., Asherson, P., Ball, D., Craig, I., Eley, T., Goldberg, D., Gray, J., Mann, A., Owen, M., & Plomin, R. (2001). Genesis: Creating a composite index of the vulnerability to anxiety and depression in a community-based sample of siblings. Twin Research, 3, 316–322.
- Shweder, R. A. (1975). How relevant is an individual difference theory of personality? Journal of Personality, 43, 455–484.
- Simpson, C. G. (1961). Principles of animal taxonomy. New York: Columbia University Press.
- Tellegen, A. (1982). Brief manual for the Differential Personality Questionnaire. Unpublished manuscript, University of Minnesota, Minneapolis.
- Tellegen, A. (1985). Structures of mood and personality and their relevance to assessing anxiety with an emphasis on self-report. In A. H. Tuma & J. D. Maser (Eds.), Anxiety and the anxiety disorders (pp. 681–706). Hillsdale, NJ: Erlbaum.
- Thomis, M. A., Vlietnick, R. F., Maes, H. H., Limkie, C. J., van Leemputte, M., Claessens, A. L., Marchal, G., & Beunen, G. P. (2000). Predictive power of individual genetic and environmental factor scores. Twin Research, 3, 99–108.
- Turkheimer, E. (1998). Heritability and biological explanation. Psychological Review, 105, 782–791.
- Turkheimer, E., & Waldron, M. (2000). Nonshared environment: A theoretical, methodological, and quantitative review. Psychological Bulletin, 126, 78–108.
- Vandenbergh, D. J., Zonderman, A. B., Wang, J., Uhl, G. R., & Costa, P. T. (1997). No association between Novelty Seeking and dopamine D4 receptor (DRD4) exon III seven repeat alleles in Baltimore Longitudinal Study of aging participants. Molecular Psychiatry, 2, 417–419.
- Vernon, P. A., Lee, D., Harris, J. A., & Jang, K. L. (1996). Genetic and environmental contributions to individual differences in alcohol expectancies. Personality and Individual Differences, 21, 183–187.
- Waller, N. G., & Shaver, P. R. (1994). The importance of nongenetic influences on romantic love styles: A twin-family study. Psychological Science, 5, 268–274.
- Watson, D., & Clark, L. A. (1997). Extraversion and its positive emotional core. In R. Hogan & J. A. Johnson (Eds.), Handbook of personality psychology (pp. 767–793). San Diego, CA: Academic Press.
- Widiger, T. A. (1993). The DSM-III-R categorical personality disorder diagnoses: A critique and alternative. Psychological Inquiry, 4, 75–90.
- Wiggins, J. S. (1979). Apsychological taxonomy of trait-descriptive terms: The interpersonal domain. Journal of Personality and Social Psychology, 37, 395–412.
- Zuckerman, M. (1991). Psychobiology of personality. Cambridge, England: University of Cambridge Press.
- Zuckerman, M. (1994). An alternative five-factor model for personality. In C. F. Halverson, Jr. & G. A. Kohnstamm (Eds.), The developing structure of temperament and personality from infancy to adulthood (pp. 53–68). Hillsdale, NJ: Lawrence Erlbaum.
- Zuckerman, M. (1995). Good and bad humors: Biochemical bases of personality and its disorders. Psychological Science, 6, 325–332.
- Zuckerman, M. (1999). Vulnerability to psychopathology: A biosocial model. Washington, DC: American Psychological Association.
- Zuckerman, M., Kuhlman, D. M., Joireman, J., Teta, P., & Kraft, M. (1993). A comparison of three structural models for personality: The Big Three, the Big Five, and the Alternative Five. Journal of Personality and Social Psychology, 65, 757–768.
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality