
Sample Genetics Of Intelligence Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
In theory, the ‘genetics of intelligence’ describes the extent to which, and manner by which, an individual’s mental abilities are influenced by his or her genes. In practice, there is a lot of debate and no universally accepted description of the genetics of intelligence. Intelligence is regarded as a complex trait, meaning that many genetic and environmental factors are thought to influence it. Nevertheless, we know little about the number or nature of genes that influence cognitive abilities. We know even less about how the expression of these genes depends on other genes or on environmental factors. To complicate matters even further, there are numerous facets of intelligence, each of which may be influenced in different ways by genes and the environment. This research paper focuses on the genetics of human intelligence and on the problems of assessing the genetic and environmental contributions to intelligence. To begin, definitions and measures of intelligence will be described. The effects of genes known to influence intelligence will then be mentioned. Finally, the degree to which variation in intelligence among humans may be genetically based will be discussed.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Intelligence Is A Multifaceted Trait
Before one can study the genetics of a trait, one has to define precisely what the trait is and how it can be measured. Once a measure has been defined for a trait, then one can begin to tackle the question of how an individual’s score may be influenced by genetic and environmental factors. Specifying an appropriate scale for intelligence is complicated by the fact that intelligence is a multifaceted trait. That is, there are several aspects of intelligence, involving reasoning, abstraction, problem solving, memory, linguistic aptitude, artistic ability, etc. Abilities in these different areas tend to be correlated to some extent within a population, which has led some authors to posit the existence of an underlying general intelligence factor, g (Spearman 1904). It is argued that the g factor directly influences many different aspects of intelligence, causing them to be correlated. Another possibility is that different aspects of intelligence interact with one another and that these interactions generate the observed correlations (Hunt 1997). For example, strength in one area of intelligence (e.g., memory) might compensate for weakness in another area (e.g., linguistic ability), causing a person’s abilities in these areas to be more similar than would be expected for uncorrelated traits. A compelling reason to believe that the various facets of intelligence are not just alternate manifestations of some underlying g factor is that they respond in different ways to aging and education. For example, Horn (1985) noted that measures of ‘fluid intelligence’ (the ability to solve new problems) generally declines with age among adults whereas ‘crystallized intelligence’ (the ability to apply known problem-solving methods to a problem at hand; see Cattell 1971) tends to increase with age and experience. A final possibility is that there are both general and specific cognitive factors that interact in a myriad of ways for different aspects of intelligences (e.g., Spearman 1904, Vernon 1979, Carroll 1993).
In most genetic studies, some variant of the Intelligence Quotient (IQ) is used as a single measure of intelligence, ignoring the multifaceted nature of intelligence (for exceptions see Cardon et al. 1992, Cardon and Fulker 1994, Pederson et al. 1994). The IQ measure grew out of studies by Alfred Binet on the ages at which children display skills of varying difficulty. Binet (1905) developed a series of age-scaled tests to assess the mental age of children. Wilhelm Stern (1916) used these tests to devise the IQ, defined as the mental age of a child divided by his or her chronological age multiplied by 100. Similar scales have been devised for adults, such as the Wechsler Adult Intelligence Scale and the Stanford-Binet Intelligence Scale. These scales have been plagued with controversy over the degree to which the measures are biased in favor of certain classes, cultures, races, or sex (Gould 1996, Schiff and Lewontin 1986, Wahlsten 1997). It is argued that there can be no completely objective measure of intelligence, since what it means to be intelligent and who is perceived to be intelligent are social constructs subject to change.
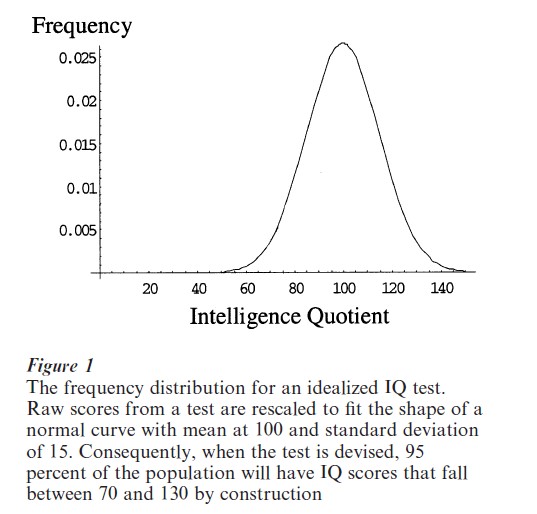
Scores on IQ tests are adjusted so that they follow a normal distribution, or a ‘bell curve,’ with a mean of 100 (Fig. 1). An indication of the importance of environmental factors on IQ is that these scales must constantly be readjusted over time, a phenomenon known as the ‘cohort effect.’ In many countries, the average IQ score has risen over time (Flynn 1987, Wahlsten 1997). For example, the average IQ score based on Ravens Progressive Matrices of 18-year old males from The Netherlands has increased from 100 in 1952 (when the test was developed and standardized) to 106 in 1962, 112.4 in 1972, and 121.1 in 1981 2. This time-span is far too short for genetic changes in the composition of the population to account for the increases. Rather, these data are evidence that changing educational and social environments can have a dramatic effect on whatever aspects of intelligence were measured by this IQ test.
2. Genes Known To Affect Intelligence
Intelligence, like any human attribute, is partly a product of our genes, simply because we would not be living human beings without these genes. However, human intelligence is partly a product of our environment, because mental impairment can be caused, for example, by exposure to toxins such as lead during development. Between these two trivial statements lies an entire spectrum of possibilities, and a heated ‘nature versus nurture’ debate has grown over the extent to which individual differences in intelligence are genetically based (Devlin et al. 1997b, Fraser 1995, Herrnstein and Murray 1994, Jensen 1969, Lewontin et al. 1984, McInerney 1996).
The human genome contains between 30,000 and 100,000 genes. Although the number of genes that affect intelligence is unknown, there is reason to believe that the number is large. Measures of intelligence, such as IQ, are most likely quantitative genetic traits influenced by hundreds or thousands of genes. Most of these genes will not be ‘intelligence’ genes, per se, but rather genes that are critical to normal development. Severe mutations in such genes would affect several traits, not just mental ability, causing a disease syndrome with multiple symptoms. These severe mutations are the most studied, in large part because they are the easiest to detect.
Geneticists have compiled over 10,000 genes and genetic disorders affecting humans into the data- base, Online Mendelian Inheritance in Man (http://www3.ncbi.nlm.nih.gov/Omim/). A search for entries describing intellectual impairment, mental retardation, or dementia turned up almost 1,000 syndromes, although many of these might not be genetically independent. Familiar examples include Down’s syndrome, Parkinson’s disease, Alzheimer’s disease, fragile X syndrome, Prader–Willi syndrome, Tay-Sachs disease, Wolf–Hirschhorn syndrome, and Rubinstein syndrome. Through the human genome project, more genetic details will be uncovered for these and other human traits and diseases.
Even in those cases where a mutation has been identified that ‘causes’ a disorder affecting intelligence, the mental development of patients carrying the mutation can be greatly improved by environmental conditions. A classic example of the interplay between genes and the environment in mental development is phenylketonuria (PKU). PKU is a recessive disease caused by a mutation at the phenylalanine hydroxylase gene (PAH) on chromosome 12. People with PKU do not produce enough of the enzyme, PAH, which oxidizes phenylalanine and converts it into tyrosine. Without this enzyme, phenylalanine levels increase to poisonous levels in the body, giving rise to a number of symptoms including mental retardation, a ‘mousy’ odor, light pigmentation, unusual posture, eczema, and epilepsy (Paine 1957). All of these manifestations of the disease may be prevented, however, by a straightforward change in the environment: a diet low in phenylalanine.
It is crucial to remember that understanding the genetics of severe mental disorders may reveal little about how intelligence develops normally and how this development depends on genetics. By analogy, understanding the environmental risk factors associated with brain damage from severe meningitis (inflammation of the tissues surrounding the brain) reveals little about environmental influences on normal brain development.
3. Population-Based Estimates Of The Genetics Of Intelligence
Although severe mutations at many genes may affect intelligence, it does not follow that the majority of differences observed among individuals within a population are due to genetic differences. In fact, genes that, when disrupted, have large effects on mental abilities may not be responsible for any of the more minor differences seen among individuals. An entirely different approach must be taken to assess the extent to which variability in a trait is based on genetic differences within a population. In animals and plants, one would raise individuals in controlled environmental conditions and measure trait differences among these individuals. Assuming that the environmental variance has been minimized in the experiment, the remaining variability is attributed to genetic differences among individuals. This controlled approach cannot be used for humans, however, and instead correlations based on family measures are used to infer the influence of genetic variation on the observed variability in a trait such as IQ.
3.1 Heritability
Heritability is a quantity used to measure the extent to which individual differences in a trait can be attributed to genetic differences. We must first define a few terms. The phenotype (P) of an individual constitutes the visible and/or measurable properties of that individual. In the following, for concreteness, we will take the phenotype to be an individual’s score on an IQ. Therefore, our discussion will focus narrowly on the heritability of IQ rather than the heritability of intelligence. Different measures of intelligence are influenced in different ways by genes and the environment and consequently have different heritabilities (Cardon et al. 1992). Nevertheless, the difficulties described below in estimating heritability apply to any measure (or measures) of intelligence in humans.
In the simplest genetic models, phenotype is imagined to be a sum of genetic and environmental effects. Specifically, the genotype (G) of an individual is the total effect of his or her genes, and the environment (E) of that individual is the total effect of all non-genetic influences (e.g., education, diet, stress) on his or her phenotype. These effects will vary among individuals, and the differences are measured by the phenotypic (VP), genotypic (VG), and environmental (VE) variance in a popul ation for a given trait. Broad-sense heritability (H0 ) measures the proportion of variance in phenotype that is attributed to genotypic variance: H0=VG /VP. This formulation assumes that the phenotype can be neatly partitioned into two parts and ignores genetic-by-environment interactions (GxE). All treatable genetic disorders, such as PKU, exhibit strong GxE, since symptoms depend on both predisposing mutations and insufficient treatment. Interactions between genes and environment may take on many forms and may well be important in the development of complex traits such as intelligence.
Broad-sense heritability estimates the degree to which differences among individuals are genetically based, but not all genetically based differences can be passed from parents to offspring. For example, whether a parent is homozygous (e.g., ++) or heterozygous (e.g., + –) at a gene is entirely a genetic attribute (broad-sense heritability is 100 percent), but this state cannot be passed on faithfully from parents to offspring. Even if both parents are heterozygous at a gene, their offspring will only be heterozygous half of the time. In other words, there are genetic attributes of an individual that depend on the particular ways in which genes combine and interact and that cannot be inferred from the additive effect (A) of each gene considered over all possible genetic backgrounds. There are two main types of genetic interactions. Dominance (D) describes interactions between the two copies of a gene (one inherited from the mother and one from the father), and epistasis describes interactions among different genes. With dominance and epistasis, knowing the phenotype of parents can sometimes provide little information about the phenotype of offspring. This point is illustrated in Fig. 2.

With genetic interactions, broad-sense heritability does not predict the extent to which offspring resemble their parents. Instead, narrow-sense heritability is used to measure how variation among individuals is influenced by genetic differences that are, on average, passed from parents to offspring. Narrow-sense heritability is defined as the fraction of phenotypic variance that can be attributed to variation in the additive effects of genes (VA): h2 = VA /VP. Narrow- sense heritability is always less than or equal to broad-sense heritability.
Neither heritability measure can be calculated directly since the genotypic (G), additive genetic (A), and environmental (E) scores for human individuals are unknowable. Instead heritability is inferred from formulae relating it to correlations among relatives. This creates an immediate problem, because the expected formulae depend strongly on the model one chooses. If one assumes that there is no epistasis, no social or cultural inheritance, no assortative mating (i.e., no correlation between parents in a trait), no sex differences, and no GxE interactions, then correlations among relatives can be derived in a straightforward manner (Table 1; see e.g., Lynch and Walsh 1998). With such a simplified model, heritability can be estimated from familial correlations. For complex traits such as IQ, however, we know that many of these assumptions are violated. Estimates in the literature for broad and narrow sense heritability differ widely, depending in large part on what set of assumptions are made.

3.2 Heritability Estimates
Bouchard and McGue (1981) summarized a large number of studies measuring the correlations between relatives in IQ scores (Table 2). Heritabilities have been estimated from these data using different models and different subsets of the data (Table 3). The heritability estimates described in Table 3 have an extremely broad range, from 0.26 to 0.72 (out of a maximum range of 0 to 1). Although a large number of models appear to be included in Table 3, most of these models share several assumptions. Consequently, the range of heritability estimates is even smaller than would be seen if models based on completely different sets of assumptions were included. For example, all estimates except the first one based on identical twins raised apart (rMZA) ignore epistasis. Genetic inter-actions are difficult to include in models because the forms of these interactions are generally not known. Without this knowledge, one cannot accurately predict correlations among relatives, just as one cannot accurately predict the winnings of the poker hands in Fig. 2 without knowing the rules of poker. Furthermore, all models ignore GxE interactions. Finally, even those models that include cultural inheritance cannot capture the intricate and diverse ways in which social structures influence IQ and the many ways in which family members can influence one another’s social context.
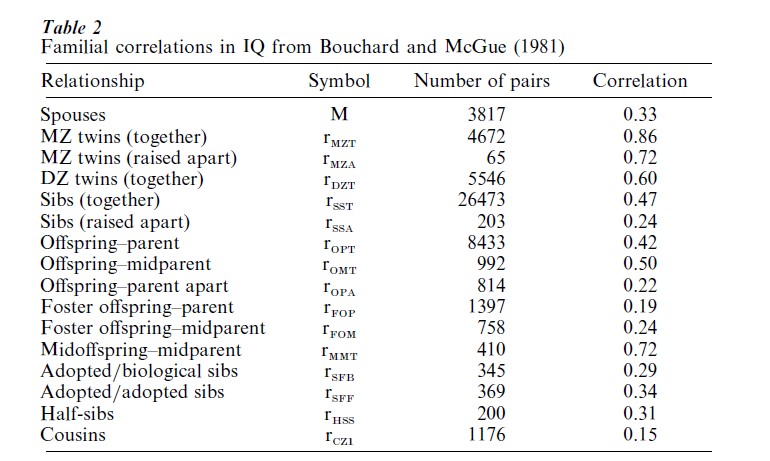
There is no consensus about the most accurate value for heritability. There are essentially two camps: those who argue that heritability lies between 0.5 and 0.8; and those who argue that heritability lies between 0.2 and 0.5. The first camp tends to focus on estimates based on twins, especially identical twins raised apart. Such estimates include dominance and epistatic inter-actions to some extent and therefore tend to measure broad-sense heritability. The correlation in IQ be- tween identical twins raised apart is indeed high (0.72), but there are several nongenetic reasons to expect that identical twins raised apart may be similar. Twins raised apart tend to be placed in similar homes (Bouchard et al. 1990). They also have a certain amount of contact before and after separation (Bouchard et al. 1990). In particular, twins share the same womb and environmental influences during gestation, including any maternal effects (Devlin et al. 1997b). Furthermore, strikingly similar twins may be more easily recognized as identical twins raised apart and may be more likely to enter twin studies. The second camp, which argues for a low heritability of IQ, tends to use a wider variety of familial correlations and more explicit models. Such models have incorporated inheritance of social and cultural influences, similarities among parents (assortative mating), selective placement of adopted children, and/or maternal effects. Although narrow-sense heritability is most often reported in these expanded studies, the difference between broad-sense and narrow-sense heritability is often small when it is measured. For example, Devlin et al. (1997a) estimate a narrow-sense heritability for IQ of 34 percent and a broad-sense heritability of 48 percent. Heritability estimates are sensitive enough to the assumptions made to warrant skepticism towards any particular value.

3.3 The Limits Of Heritability
Despite the fact that a lot of effort and scores of publications have been devoted to estimating the heritability of IQ, heritability provides very limited information of relevance to human society. Heritability is a descriptor of the variability currently within a population and how this variability is partitioned into genetic and nongenetic influences. Even with an estimate of heritability, we know nothing about the reasons for the IQ of any given individual. Furthermore, heritability depends entirely on the set of environments in which it is measured. If one changes the environment, heritability may change in unpredictable ways, both because the phenotypic variance depends on the variance in environmental effects and because genotypic effects may depend on the environment (GxE). One cannot say on the basis of heritability whether or not a trait will change in response to an environmental manipulation (e.g., improved parental support and training, dietary supplements to pregnant women, different methods of schooling).
If heritability for IQ were 100 percent, one might be tempted to conclude that environmental factors are irrelevant and that there is little point in embarking upon social programs to improve the environment. This conclusion would, however, be entirely wrong. Consider the following fictitious example: imagine that there are only two types of people in equal frequencies, those with PKU and those without it. Imagine that no treatment is known, and that there are no environmental /cultural /social /maternal effects on IQ. Let the IQ of those without PKU equal 100, and the IQ of those with PKU equal some number less than 100. Under this scenario, 100 percent of the variability in IQ within the population is genetic (H0=1). If the link between dietary phenylalanine and mental deficits in people with PKU is then discovered, and if people with PKU start controlling their diets, the average IQ would rise towards 100, despite the fact that, originally, there was only genetic and no environmental variation within the population. In short, heritability says nothing about how IQ might change in a new environmental context.
Estimating heritability will not help us know how to improve a trait, be it IQ or health or happiness. Detailed studies about how changes in social, educational, medical, and economic opportunities affect intelligence are required to determine what programs would be effective in permitting individuals to attain their highest potential.
4. Race, Gender, And IQ
Nowhere has the debate over the genetics of intelligence been fiercer than in the literature discussing differences in IQ between genders or among races (see, for example, the debate over The Bell Curve (by Allen et al. 1996, Devlin et al. 1997b, Fraser 1995, Herrnstein and Murray 1994, McInerney 1996). In those cases where a mean difference in IQ is observed between two groups, it is very difficult to determine the root causes of the IQ difference. One problem is the inherent bias in IQ tests favoring those individuals that come from the same cultural milieu and social class as the scientists that developed the tests (for examples, see Schiff and Lewontin 1986, pp. 21–3). A further problem is that the basis of differences among groups need bear no relationship to the basis of differences within groups. Even if IQ were entirely genetically heritable within groups, IQ differences among them may be entirely cultural or environmental. For example, the difference in average IQ score between a group of children exposed in utero to high levels of alcohol and a control group would be primarily environmental in origin (due to alcohol exposure), regardless of the heritability of IQ in either group.
Perhaps the greatest hurdle to understanding the root causes of IQ differences, however, is that it is difficult to eradicate the many socioeconomic differences faced by individuals of different ethniciticies, cultures, or genders. Nevertheless, transracial adoption studies have shown that differences in IQ among racial groups largely disappear when children are adopted into caucasian families, presumably because the children are then reared in ‘the culture of the tests’ (Weinberg et al. 1992). Furthermore, those programs that have enriched the educational environment of disadvantaged children have had great success in boosting their mean IQ score. For example, Wahlsten (1997) reviews two studies of children from disadvantaged homes with few financial and educational resources; a majority of these children were African American. In the studies, children were randomly assigned to a group receiving no additional educational resources and a second group receiving educational day care prior to public schooling. The average IQ score at four years old was 13.7 and 12.1 points higher in the two studies for the children receiving additional educational resources. Considering the average IQ gap between African Americans and Caucasians in the USA is about 15 points (Devlin et al. 1997b), these studies suggest that most if not all of the differences could be eradicated by providing quality educational opportunities for all children.
5. Concluding Remarks
Nearly the entire sequence of the human genome is now available. This genetic database will make it easier to find genes associated with human traits, including aspects of intelligence. It will be essential to remember, however, that genes do not cause intelligence. The developmental process that led to who we are today is a complex one involving the expression and repression of tens of thousands of genes in a manner that both depends on environmental conditions and alters these environmental conditions. The specific form of a gene that may enhance IQ in one family in one set of environments may be irrelevant in other families and in other environments. If future genetic studies of intelligence are able to identify genes that enhance (or reduce) IQ under certain circumstances, then we may be able to mimic (or counter) the effects of these genes. Attempts to improve the educational and nutritional environments of children are, however, much more promising and immediate. Yet we should remember that linear, one-dimensional scales such as IQ mislead us into thinking that there are better and worse intellects. In reality, human thought is much more complex and multi-dimensional, which we must keep in mind if we are to preserve and encourage the rich intellectual diversity that characterizes human society.
Bibliography:
- Allen A, Anderson B, Andrews L, Beckwith J, Bowman J et al. 1996 The Bell Curve: statement by the NIH-DOE Joint Working Group on the ethical, legal, and social implications of human genome research. American Journal of Human Genetics 59: 487–8
- Binet A A S T 1905 Methodes nouvelles pour le diagnostic du niveau intellectuel des anormaux [New methods for diagnosing the intellectual level of abnormal persons]. Annee Psychologique 11: 191–336
- Bouchard T J, Lykken D T, McGue M, Segal N L, Tellegen A 1990 Sources of human psychological differences: the Minnesota study of twins reared apart. Science 250: 223–28
- Bouchard T J, McGue M 1981 Familial studies of intelligence: a review. Science 212: 1055–9
- Cardon L R, Fulker D W 1994 A model of developmental change in hierarchical phenotypes with application to specific cognitive abilities. Behavior Genetics 24: 1–16
- Cardon L R, Fulker D W, DeFries J C 1992 Multivariate genetic analysis of specific cognitive abilities in the Colorado adoption project at age 7. Intelligence 16: 383–400
- Carroll J B 1993 Human Cognitive Abilities. Cambridge University Press, Cambridge, UK
- Cattell R B 1971 Abilities: Their Structure, Growth, and Action. Houghton Mifflin, Boston, MA
- Cavalli-Sforza L L, Feldman M W 1978 Dynamics and statistics of traits under the influence of cultural transmission. In: Morton N E, Chung C S (eds.) Genetic Epidemiology. Academic Press, New York, pp. 133–43
- Cloninger C R, Rice J, Reich T 1979a Multifactorial inheritance with cultural transmission and assortative mating. II. A general model of combined polygenic and cultural inheritance. American Journal of Human Genetics 31: 176–98
- Cloninger C R, Rice J, Reich T 1979b Multifactorial inheritance with cultural transmission and assortative mating. III. Family structure and the analysis of separation experiments. American Journal of Human Genetics 31: 366–88
- Devlin B, Daniels M, Roeder K 1997a The heritability of IQ. Nature 388: 468–71
- Devlin B, Fienberg S E, Resnick D P, Roeder K 1997b Intelligence, Genes, and Success: Scientists Respond to The Bell Cur e. Springer, New York
- Feldman M W, Cavalli-Sforza L L 1979 Aspects of variance and covariance analysis with cultural inheritance. Theoretical Population Biology 15: 276–307
- Flynn J R 1987 Massive IQ gains in 14 nations: what IQ tests really measure. Psychological Bulletin 101: 171–91
- Fraser S 1995 The Bell Cur e Wars: Race, Intelligence, and the Future of America. BasicBooks, New York
- Gould S J 1996 The Mismeasure of Man. Norton, New York
- Herrnstein R J, Murray C 1994 The Bell Cur e: Intelligence and Class Structure in American Life. Free Press, New York
- Horn J L 1985 Remodelling old models of intelligence. In: Wolman B B (ed.) Handbook of Intelligence: Theories, Measurements, and Applications. Wiley, New York pp. 267–300
- Hunt E 1997 The concept and utility of intelligence. In: Devlin B, Fienberg S E, Resnick D P, Roeder K (eds.) Intelligence, Genes, and Success: Scientists Respond to The Bell Cur e. Springer, New York, pp. 157–76
- Jensen A R 1969 How much can we boost IQ and scholastic achievement. Harvard Educational Review 39: 1–123
- Lewontin R C, Rose S P R, Kamin L J 1984 Not in Our Genes: Biology, Ideology, and Human Nature, 1st edn. Pantheon Books, New York
- Lynch M, Walsh B 1998 Genetics and Analysis of Quantitative Traits. Sinauer, Sunderland, MA
- Martin N G, Eaves L J, Heath A C, Jardine R, Feingold L M et al 1986 Transmission of social attitudes. Proceedings of the National Academy of Sciences of the United States of America 83: 4364–8
- McInerney J D 1996 Why biological literacy matters: a review of the commentaries related to The Bell Curve: intelligence and class structure in American life. The Quarterly Review of Biology 71: 81–96
- Morton N E, Rao D C 1978 Quantitative inheritance in man. Yearbook of Physical Anthropology 21: 12–41
- Otto S P Feldman M W Christiansen F B 1994 Genetic and cultural transmission of continuous traits. In: Morrison Institute Working Paper. Stanford University, Stanford, CA, pp. 1–91
- Paine R S 1957 The variability in manifestations of untreated patients with phenylketonuria (phenylpyruvic aciduria). Pediatrics 20: 290–302
- Pedersen N L, Plomin R, McClearn G E 1994 Is there G beyond g? (Is there genetic influence on specific cognitive abilities independent of genetic influence on general cognitive ability?) Intelligence 18: 133–43
- Plomin R, Loehlin J C 1989 Direct and indirect IQ heritability estimates: a puzzle. Behavior Genetics 19: 331–42
- Rao D C, Morton N E, Cloninger C R 1979 Path analysis under generalized assortative mating. I. Theory. Genetical Research of Cambridge 33: 175–88
- Rao D C, Morton N E, Yee S 1976 Resolution of cultural and biological inheritance by path analysis. American Journal of Human Genetics 28: 228–42
- Rice J, Cloninger C R, Reich T 1978 Multifactorial inheritance with cultural transmission and assortative mating. I. Description and basic properties of the unitary models. American Journal of Human Genetics 30: 618–43
- Schiff M, Lewontin R C 1986 Education and Class: The IrRelevance of IQ Genetic Studies. Oxford University Press, Oxford, UK
- Spearman C 1904 General intelligence, objectively determined and measured. American Journal of Psychology 15: 201–93
- Stern W 1916 Der Intelligenz Quotient vals Masz der kindlichen Intelligenz, inbesondere der Unternormal [The Intelligence Quotient as a measure of children’s intelligence, especially for those below normal]. Zeitschrift fur angewandte Psychologie 11: 1–18
- Vernon P E 1979 Intelligence: Heredity and Environment. Freeman, San Francisco
- Wahlsten D 1997 The malleability of intelligence is not constrained by heritability. In: Devlin B, Fienberg S E, Resnick S E, Roeder K (eds.) Intelligence, Genes, and Success: Scientists Respond to The Bell Curve. Springer, New York, pp. 71–87
- Weinberg R A, Scarr S, Waldman I D 1992 The Minnesota transracial adoption study: a follow-up of IQ test performance at adolescence. Intelligence 16: 117–35
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality