
Sample Primate Socioecology Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our research paper writing service for professional assistance. We offer high-quality assignments for reasonable rates.
Sociality is the primary behavioral adaptation of the anthropoid primates, the monkeys and apes. All of the anthropoid primates, except orangutans, spend most of their lives in stable groups of familiar individuals. However, the social organization among the anthropoid primates encompasses great diversity, ranging from the pair-bonded groups of gibbons and owl monkeys, to the polyandrous groups of tamarins; onemale groups of howlers and blue monkeys; multimale, multifemale groups of cebus monkeys and baboons; and fission-fusion communities of chimpanzees and spider monkeys. This great diversity in social organization shapes the Behavioral options available to males and females and profoundly influences the evolution of social Behavior and reproductive strategies in both sexes.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. The Evolution Of Social Organization
The great diversity in primate social organization has prompted researchers to ask a number of related questions: Why do primates live in groups? How should groups be structured? How big should groups be? These questions were first raised in the 1960s and 1970s by primatologists who conducted comparative analyses searching for broad ecological correlates of social organization (reviewed by Janson 2000). These comparative efforts yielded some consistent associations between diet, home range size, group size, body weight, and group composition. But the analyses were unsatisfying because they could not account for much of the observed variation in social organization among primates. Moreover, while these studies revealed a number of robust associations among various ecological variables, they did not produce much insight about the causal processes underlying these correlations. Finally, these comparative studies did not consider the possibility that some of the similarities between species were the result of common phylogenetic history, not independent adaptations to ecological conditions, a problem that has become a central methodological concern in comparative studies in Behavioral ecology.
In the 1980s researchers returned to these questions, equipped with the theoretical and methodological tools of contemporary Behavioral ecology (Krebs and Davies 1993). Behavioral ecologists try to explain how evolutionary processes shape Behavior in particular ecological contexts. Sociality is presumed to reflect a dynamic balance between the advantages and disadvantages of living in close proximity to conspecifics. Sociality is beneficial because animals who live in social groups are better able to acquire and control resources or less vulnerable to predators. On the other hand, sociality is disadvantageous because groupliving animals may face more competition over access to food, become more vulnerable to disease, and become more susceptible to cuckoldry, infanticide, and cannibalism. The size and demographic composition of the groups that we see in nature reflects a compromise between the costs and benefits of sociality for individuals who occupy particular ecological niches.
1.1 Why Do Primates Live In Groups?
For primates, the primary advantages of sociality are thought to be enhanced access to resources, reduced vulnerability to predation, or some combination of both these factors. Many primates feed on foods, such as fruit, that occur in patches large enough to feed several individuals at the same time. Taking note of this fact, Wrangham (1980) suggested that primates aggregate because groups are more successful in defending access to resources than lone individuals, an argument that has also been applied in other taxa. Thus, the potential for between-group competition over resources favors sociality.
van Schaik (1983, 1989) questioned Wrangham’s explanation of the evolution of sociality, arguing that the benefits accrued in between-group competition would be offset by the costs incurred from increased within-group competition. Instead, van Schaik argued, grouping evolved as a defense against predators. Primates are preyed upon by a variety of predators, including leopards, raptors, snakes, and other primates. Grouping provides safety from predators for three reasons. First, levels of vigilance are higher because there are more individuals scanning for predators. Second, groups are more effective in actively mobbing or deterring predators. Third, any single individual is less likely to be caught in a group if predators strike at random. Thus, primates aggregate to obtain protection from predators, and within-group competition for food or safe positions is an unavoidable side effect of sociality.
Initial efforts to evaluate the relative importance of between-group competition and predation among primates provided some support for both models (Janson 2000). Thus, the between-group competition model was supported by evidence that hostile intergroup interactions are common among primates, and larger groups are typically able to supplant smaller groups in intergroup encounters. In some populations, large groups are better able to defend higher quality home ranges than smaller groups. However, within-group competition seems to reduce foraging efficiency consistently in gregarious species (Janson 1992).
The predation model was supported by evidence that group sizes are reduced when predators are absent and that members of large groups spend less time scanning for predators and detect predators sooner than members of small groups do (Janson 1992). Nonetheless, vigilance levels do not decline monotonically as group size increases (Janson 2000), suggesting that groups are larger than needed for predator defense.
1.2 The Nature Of Competition And Female Social Relationships
As researchers continued to debate the role of resource competition and predation in the evolution of sociality, they also began to think more carefully about the effects of competition on primate females. The distribution of food resources determines the distribution of females, and this in turn determines the distribution of males. Females come first in this scenario because female fitness depends mainly on their nutritional status (Wrangham 1980), while male fitness depends primarily on their ability to obtain access to fertile females.
Wrangham (1980) hypothesized that variation in the extent of intergroup competition would produce two distinct kinds of social arrangements. When resources were clumped and abundant enough to feed more than one individual, females would benefit from banding together to collectively defend access to them. Coalitionary alliances among kin are expected to be more beneficial than alliances among nonkin because kin share a common genetic interest (Hamilton 1964); thus females were expected to form nepotistic alliances. Female philopatry preserves matrilineal networks, and strong affiliate bonds strengthen alliances. Thus, female-bonded kin groups are expected to be associated with between group competition over resources. On the other hand, if resources are not clumped or occur in patches that are too small to feed several individuals, then the collective defense of resources will not be profitable. Nepotistic alliances will not enhance females’ access to resources. In this situation, selective pressures favoring female philopatry and strong female bonds will be relaxed. When there is little between-group competition, non-female bonded groups are expected to occur. Wrangham linked dietary specializations to social organization, arguing that fruit occurs in discrete patches that are often large enough to make collective defense profitable. He predicted that female-bonded groups would be mainly frugivorous.
Although it was rather easy to classify primate species as female-bonded or non-female-bonded, a number of empirical and theoretical objections to Wrangham’s model were raised. However, all subsequent efforts to link ecology and social Behavior have relied on Wrangham’s insight that resource competition among females drives the evolution of social organization among primates.
van Schaik (1989) considered how the distribution of resources influences the nature of competition among females. Scramble competition occurs when individuals cannot effectively monopolize access to resources, and consequently do not compete over them directly. Contest competition occurs when resources are limited and can be monopolized profitably, generating direct confrontations over access to them. van Schaik suggested that within group contest competition (WGC), within group scramble competition (WGS), and between group contest competition (BGC) would have different effects on social organization (Table 1). (Between group scramble competition is assumed to be an automatic consequence of population density, but is not expected to influence social relationships directly).
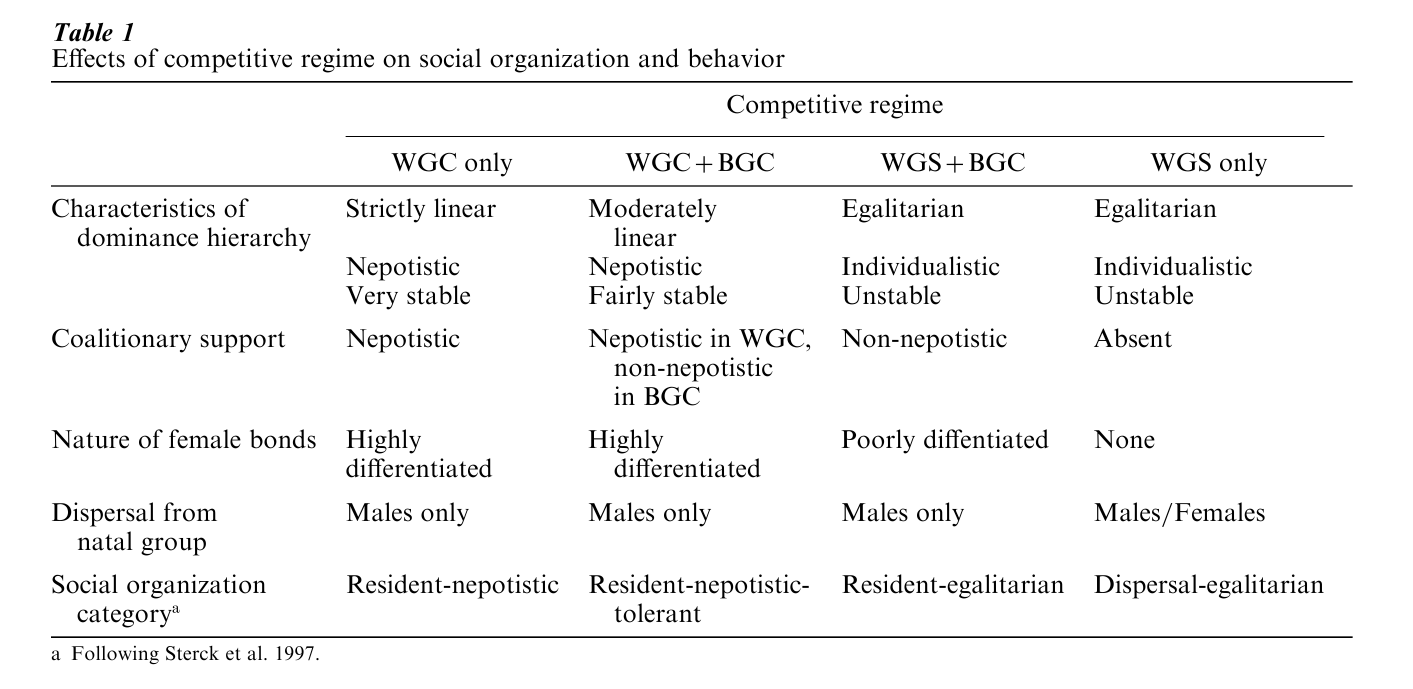
When females compete over access to resources with group members, access to resources will be a function of individual dominance rank Van Schaik predicted that WGC would be associated with strict linear dominance hierarchies. Since dominance rank influences female access to resources and ultimately affects their fitness, females will provide coalitionary support to their kin in order to enhance their dominance rank. This will lead to matrilineal dominance hierarchies and create strong selective pressures favoring female philopatry. Nepotistic support will also enhance the stability of dominance hierarchies by dampening the effects of temporary variations in individual fighting ability. Affiliative interactions with coalition partners will reinforce alliances, so females are expected to form close and well-differentiated social relationships.
When WGS is the primary form of resources competition, female reproductive success will depend on the size of the female’s social group, not her own rank. This means that there will be little incentive to aid relatives in disputes with other group members, and dominance hierarchies will not be nepotistic or strictly linear. If WGS is combined with strong BGC, females will rely on coalitionary support in intergroup encounters from all group members. Because females benefit more from supporting kin than nonkin, female philopatry will be favored. Since females’ fitness depends on collective group action, females are expected to cultivate social ties with all other group members. Thus, females are expected to form relatively poorly differentiated social relationships.
If WGS is associated with weak BGC, then selective pressures favoring female philopatry will be relaxed. Female residence in social groups should reflect the balance between the benefits of living in groups of particular sizes and the costs of emigration from groups of suboptimal size.
1.3 Testing Socioecological Models In The Field
In the 1990s, many researchers set to work testing the predictions derived from socioecological models. This body of work confirmed many of the predictions derived from socioecological models. For example, Sterck et al. (1997) showed that coalitionary support, linear dominance hierarchies, and female philopatry are linked in a sample of 27 primate taxa. Others have compared closely related species, that differ in key features of their ecology. For example:
(a) Mitchell et al. (1991) compared two closely related species of squirrel monkeys in Costa Rica and Peru that had similar diets, formed groups of similar sizes, and experienced similarly high levels of predation. The two species differed mainly in the extent of within-group competition over access to food resources. As predicted by socioecological models, the species that experienced high within-group competition also formed stable, linear dominance hierarchies, kin-based coalitions, and exhibited strict female philopatry. In the species that experienced low levels of within-group competition, a dominance hierarchy was not detected among females, female coalitions were not observed, and females sometimes left their natal groups.
(b) Barton et al. (1996) compared the Behavior and ecology of baboons at two sites. In Laikipia, Kenya, where within-group competition for food and predation pressures were high, groups tended to be large, females formed strong bonds with one another, and females rarely transferred from their natal groups. In the Drakensburg Mountains of South Africa, food was more evenly distributed and predation pressure was relaxed. There, groups were typically smaller, bonds among females were weaker, female dispersal was more common, and bonds between females and males were strengthened.
(c) Isbell and her colleagues have examined the relative importance of phylogeny and ecology in shaping the Behavior of female monkeys in two closely related species, patas (Erythrocebus patas) and vervets (Cercopithecus aethiops). If phylogeny is the primary factor influencing social organization, then patas and vervets should be very similar. However, if ecological factors are crucial, then patas and vervets should differ because their diets differ. Patas rely more heavily than vervets on arthropods and other food items that require little handling time and are consumed very quickly. On the other hand, vervets rely more heavily than patas on foods, such as gums, seeds, and fruits, that have longer processing times and take longer to eat. Patas have less linear dominance hierarchies than vervets do, and dominance rank has a less pronounced effect on nonagonistic Behavior tightly linked to ecology (such as ranging, diet, and activity budgets) among patas than among vervets (Isbell et al. 1999).
(d) Koenig et al. (1998) compared two populations of Hanuman langurs (Presbytis entellus) living at different sites in India. The langurs of Ramnagar relied heavily on just three food plants. These plants were low in abundance and clumped in their distribution, conditions that are expected to lead to contest competition. The langurs of Kanha relied on a larger array of plant foods, and one of their most highly preferred foods is superabundant, conditions that are expected to lead to scramble competition. The langurs of Ramnagar behave much as expected when WGC is strong: they have linear dominance hierarchies and females are philopatric. The langurs of Kanha have poorly developed dominance hierarchies and females frequently emigrate from their natal groups.
1.4 Distribution Of Males
In socioecological models, males generally play a secondary role. Ecological factors shape the distribution of females, and males go where females are (Kappeler 2000). Social factors are generally thought to be more important than ecological factors in determining the distribution of males because instrasexual competition over access to females contributes more to variation in male reproductive success than intrasexual competition over access to food. When females live in groups, they become a defensible resource for males. The more females there are, the more difficult it is for a single male to monopolize them. This difficulty is compounded if females are widely dispersed or if females become receptive simultaneously. Comparative analyses indicate that the number of males in primate groups is generally correlated with the number of females present and with the extent of synchrony in female receptivity (Mitani et al. 1996, Nunn 1999).
The availability of females explains some, but not all of the variation in the number of males in social groups. This raises the possibility that the distribution of males is also influenced by other factors, including ecological pressures. Comparative analyses suggest that the distribution of males in social groups is directly linked to predation pressure. Hill and Lee (1998) conducted a systematic survey of the Old World monkeys in an effort to assess the effects of predation risk on the distribution of males. They found that predation risk consistently influenced group size, as groups that faced the highest risks of predator attacks lived in the largest groups. In single-male groups, high predation risk was associated with an increase in the number of females, but in multimale groups predation risk influenced the number of males in social groups, but not the number of females. Thus, the distribution of males may be a joint product of the distribution of females and the intensity of predation pressures.
2. Beyond Socioecology
Socioecological models of primate social organization gained considerable currency in the 1990s. However, in the last few years, socioecological models of social organization have been subjected to a new set of empirical and theoretical challenges (Janson 2000). Not all the observed variation in social organization and Behavior fit predictions derived from socioecological models. For example the comparison of squirrel monkeys by Mitchell et al. was complicated by subsequent observations of squirrel monkeys in Suriname (Boinski 1999). There, monkeys face intense WGC, but females have weak bonds with one another and do not form coalitions with other females. Comparative evidence indicates that males generally go where females are, but it has been surprisingly difficult to explain why individual males move from one group to another. Male baboons do not always enhance their access to females when they emigrate (Altmann 2000).
The role of environment in shaping social organization was challenged by new comparative studies which took phylogeny explicitly into account (di Fiore and Rendall 1994). Selected pairwise comparisons of closely related primate species provide support for socioecological models, but these comparisons may not be representative. Most of the characteristics associated with WGC, including linear dominance hierarchies, nepotistic alliances, and female philopatry are found in almost all of the extant Cerocipthecine species, even though they now occupy an extremely diverse range of habitats. Di Fiore and Rendall’s analysis indicates that there is considerable inertia in social evolution. The social systems categorized by socioecologists may represent different peaks in the adaptive landscape (Pope 2000). If so, this seriously constrains the possible paths for change in social evolution.
Socioecological models assume that ecological factors play a primary role in shaping social organization, but a strong case can be made that demographic and social factors may also shape social systems. A growing body of evidence suggests that the risk of infanticide may affect primate social systems (van Schaik 2000). There is compelling evidence that infanticide is an evolved reproductive strategy in many animal taxa. In many primate species, males selectively kill infants whose deaths enhance their own reproductive prospects (van Schaik and Janson 2000). The presence of multiple males in social groups throughout the year may be linked to infanticidal risks faced by females (van Shaik 2000).
In summary, present evidence suggests that a broad range of ecological, historical, demographic, and social factors influence the evolution of primate groups. It was probably naıve to expect simple models to explain the full range of variation in primate social organization and Behavior. It is, however, essential to begin with simple models before constructing more elaborate ones. The task now is to elaborate and extend socioecological models to accommodate new data and ideas about why primates live in groups.
Bibliography:
- Altmann J 2000 Models of outcome and process: Predicting the number of males in primate groups. In: Kappeler P M (ed.) Primate Males. Cambridge University Press, Cambridge, UK
- Barton R A, Byrne R W, Whiten A 1996 Ecology, feeding competition, and social structure in baboons. Behavioral Ecology and Sociobiology 38: 321–9
- Boinski S 1999 The social organizations of squirrel monkeys: Implications for ecological models of social evolution. Evolutionary Anthropology: Issues, News, and Reviews 9: 101–12
- di Fiore A, Rendall D 1994 Evolution of social organization: A reappraisal for primates by using phylogenetic methods. Proceedings of the National Academy of Science USA 91: 9941–95
- Hamilton W D 1964 The genetical evolution of social behavior I, II. Journal of Theoretical Biology 7: 1–52
- Hill R A, Lee P C 1998 Predation risk as an influence on group size in cercopithecoid primates: implications for social structure. J. Zool. Lond. 245 (part 4): 447–56
- Isbell L A, Pruetz J D, Lewis M, Young T P 1999 Rank differences in ecological Behavior: A comparative study of patas monkeys (Erythrocebus patas) and vervets (Cercopi-thecus aethiops). International Journal of Primatology 20: 257–72
- Janson C H 1992 Evolutionary ecology of primate social structure. In: Smith E A, Winterhalder B (eds.) Evolutionary Ecology and Human Behavior. Aldine de Gruyter, New York, NY
- Janson C H 2000 Primate socioecology: The end of a golden era. Evolutionary Anthropology: Issues, News, and Reviews 9: 73–86
- Kappeler P M 2000 Primate males: history and theory. In: Kappeler P M (ed.) Primate Males. Cambridge University Press, Cambridge, UK
- Koenig A, Beise J, Chalise M K, Ganzhorn J U 1998 When females should contest for food—testing hypotheses about resource density, distribution, size, and quality with Hanuman langurs (Presbytis entellus). Behavioral Ecology and Sociobiology 42: 225–237
- Krebs J R, Davies N B 1993 An Introduction to Behavioural Ecology, 3rd edn. Blackwell, Oxford, UK
- Mitchell C L, Boinski S, van Schaik C P 1991 Competitive regimes and female bonding in two species of squirrel monkeys (Saimiri oerstedi and S. sciureus). Behavioral Ecology and Sociobiology 28: 55–60
- Mitani J C, Gros-Louis J, Manson J H 1996 Number of males in primate groups: Comparative tests of competing hypotheses. American Journal of Primatology 38: 315–32
- Nunn C L 1999 The number of males in primate social groups: A comparative test of the socioecological model. Behavioral Ecology and Sociobiology 46: 1–13
- Pope T R 2000 The evolution of male philopatry in neotropical monkeys. In: Kappeler P M (ed.) Primate Males. Cambridge University Press, Cambridge, UK
- Sterck E H M, Watts D P, van Schaik C P 1997 The evolution of female social relationships in nonhuman primates. Behavioral Ecology and Sociobiology 41: 291–309
- van Schaik C P 1983 Why are diurnal primates living in groups? Behaviour 87: 120–44
- van Schaik C P 1989 The ecology of social relationships amongst female primates. In: Standon V, Foley R A (eds.) Comparative Socioecology: the Behavioural Ecology of Humans and Other Mammals. Blackwell, Oxford, UK
- van Schaik C P 2000 Social counterstrategies against infanticide by males in primates and other mammals. In: Kappeler P M (ed.) Primate Males. Cambridge University Press, Cambridge, UK
- van Schaik C P, Janson C H (eds.) 2000 Infanticide by Males and its Implications. Cambridge University Press, Cambridge, UK
- Wrangham R W 1980 An ecological model of female-bonded primate groups. Behaviour 75: 262–300
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality