
Sample Population Dynamics Theorems Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. iResearchNet offers academic assignment help for students all over the world: writing from scratch, editing, proofreading, problem solving, from essays to dissertations, from humanities to STEM. We offer full confidentiality, safe payment, originality, and money-back guarantee. Secure your academic success with our risk-free services.
1. Extinction, Stable Size, Or Explosion?
Historically, probabilistic population theories originate from the extinction problem: suppose you know the probabilities of begetting 0, 1, 2, 3,… children. Now determine the probability that a population of a known number of individuals will eventually die out. Already Malthus had noted that a rapid increase of the whole population could occur together with frequent extinction of the separate families constituting the population. Rephrased in the terminology of evolutionary biology, frequent extinction of species does not contradict a rapid growth of the biomass of the world.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
The mathematical explanation of this apparent paradox is due to Bienayme (1845) and independently, thirty years later and only in parts correctly, to Galton and Watson, who were later to lend their names to the simplest, generation-counting version of the so called branching processes that provide the mathematical structure underlying most stochastic population dynamics. Indeed, if pk denotes the probability of giving birth to k children, k = 0, 1, 2, 3,… then the extinction probability of a population stemming from one ancestor will be the smallest nonnegative solution q of the equation
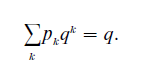
If we disregard the trivial case p1 =1, q will equal 1 if and only if the mean number of children
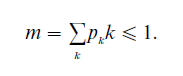
If m > 1, then the population growth rate per generation will be m, and even though q must be < 1 it can still be pretty large: it is easy to formulate demographically or biologically plausible cases where, say, m exceeds 2 and q is still around 0.8.
If a stochastic population does not die out, it can be proved very generally that it must grow beyond all limits (see Jagers 1991). This is a consequence of stochasticity and the asymmetry between extinction and big sizes: however big the population, there remains an albeit small but still positive risk of extinction, whereas an extinct population cannot be resurrected. This is in contradiction to deterministic population theories which allow population sizes to be stable, cf. Keyfitz 1968.
A finite world can certainly not sustain infinite growth. One can therefore argue that probabilistic population dynamics show that all real populations will die out sooner or later. But before that happens there may be a long period of growth.
2. Population Growth
To describe population growth in real time rather than generation-wise, a more sophisticated description of individual life and reproduction is needed. Probabilistic population dynamics is indi idual based in the sense that it starts from such a description. Deterministic theories, on the other hand, often start directly from an assumed behavior of the population as a whole. Demographic theory is often pseudo probabilistic in that it uses probabilistic notions but makes conclusions only about the expected behavior of the population as a whole. (This is the reason why there exist deterministic populations with stationary sizes: if the underlying stochastic population is critical in the sense that on average each individual is replaced by one child, then the expected total population size will be stationary, whereas the population itself will die out sooner or later.)
The most general probabilistic formulation of individual life goes as follows: at birth an individual inherits a type from her mother. The type determines her survival distribution as well as the distribution of age at childbearing. It also determines the types of the children she will give birth to in her turn. (Think of the type as a genotype, or a genotype combined with the social and biological conditions of life.)
In the supercritical case, where there is a positive chance of nonextinction, the probability laws of survival and childbearing determine a positive Malthusian parameter, which gives the rate of exponential growth. In the theory of branching processes this rate is usually denoted by α, so that the population will grow like eαt, as time t passes. In deterministic theory the conventional notation is often r instead.
Even though exponential growth appears to be a consequence of the independence assumed (tacitly or explicitly) between the reproduction of different individuals, it is interesting to note that such growth has recently been proved also to occur in many cases where individual reproduction depends upon the population as a whole, e.g., its size. For such results in the case of simple Galton-Watson branching processes see Klebaner (1984).
In cases where the reproduction approaches criticality as the population grows, non-Malthusian growth can occur nevertheless, e.g., in the form of linear growth (Klebaner 1994).
3. Stable Population Theory
In contrast to population size, the composition of growing populations tends to stabilize. The description of this composition, or rather the various stable compositions pertaining to different reproduction and survival laws, is the object of stable population theory.
The most well known aspect of stable population composition is the stable age distribution, first described by Euler in 1760 and since then rediscovered repeatedly: if the Malthusian parameter is α > 0 and all individuals have the same survival function k, then the age of an individual sampled at random in an old population will have the density
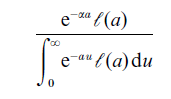
at age a, exactly as in deterministic theory. This is, of course, a law-of-large-numbers effect; we are considering ever larger populations.
If, as in the general setup mentioned above, there are several types of individuals, each with its own life span distribution, then the situation will be more complex. First of all, the type distribution will converge to a stable type law, determined by an eigen-measure property. Then the survival function l above will have to be replaced by an average of all the different survival functions weighted by the stable type distribution.
But the stable composition describes many other aspects of a balanced growing population besides ages. For example the probability of being first-born can be calculated as well as other aspects of family structure. In modern evolutionary biology the genealogy of individuals and, say, the mutational history plays an important role. In an old population this is described by a stable, typical ancestry, which can be used to determine time back to some event like divergence between species.
Bibliography:
- Athreya K B, Ney P 1972 Branching Processes. Springer-Verlag, Berlin
- Harris T 1963 The Theory of Branching Processes. SpringerVerlag, Berlin
- Jagers P 1975 Branching Processes with Biological Applications. Wiley, Chichester
- Jagers P 1991 The growth and stabilization of populations. Statistical Science 6: 269–83
- Keyfitz N 1968 Introduction to the Mathematics of Population. Addison-Wesley, Reading, MA
- Klebaner F C 1984 Geometric rate of growth in population size dependent branching processes. Journal of Applied Probability 21: 40–9
- Klebaner F C 1994 Asymptotic behaviour of Markov population processes with asymptotically linear rate of change. Journal of Applied Probability 31: 614–25
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality