
View sample tactile information processing in the primate brain research paper. Browse research paper examples for more inspiration. If you need a psychology research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our writing service for professional assistance. We offer high-quality assignments for reasonable rates.
In this research paper, we discuss the neural mechanisms of tactile perception. In each of the sensory systems, information about the external world is analyzed into separate processing streams. In the somatosensory system, that division begins at the very first stage of sensory processing. Nociceptors, thermoreceptors, proprioceptors, and cutaneous mechanoreceptors transduce different stimulus features and channel their information into separate, parallel streams.We focus here on the four cutaneous mechanoreceptors that are responsible for tactile perception. Evidence from psychophysical and neurophysiological studies, reviewed here, suggests that they serve distinctly separate functions. We first describe the four mechanoreceptor types and discuss their roles in tactile perception. Then we discuss the anatomy and functions of the central pathways.We end with a discussion of the role of attention in tactile information processing. Throughout the paper we focus on the processing of information about surface features and object form.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
Peripheral Neural Mechanisms of Tactile Perception
Mechanoreception
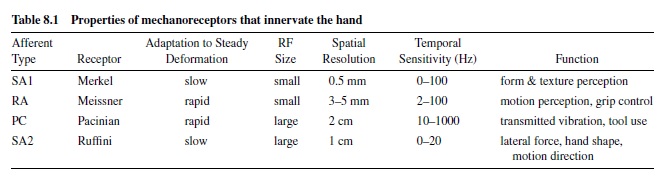
Tactile perception is based on four cutaneous mechanoreceptive afferent neuron types: slowly adapting type 1 (SA1), rapidly adapting (RA), Pacinian (PC), and slowly adapting type 2 (SA2) afferents (see Table 8.1). Two of the four, the RA and PC afferents, respond only to skin motion; they are classed as rapidly adapting because they respond transiently to sudden indentation. The other two, the SA1 and SA2 afferents, are classed as slowly adapting because they respond to sustained skin deformation with a sustained discharge that declines slowly, although they (particularly SA1 afferents) are more sensitive to skin movement than to static deformation. The neural response properties of these cutaneous afferents have been studied extensively in both human and nonhuman primates, and—except for the SA2 afferents, which are not found in nonhuman primates—there are no interspecies differences. We use the terms SA1, SA2, RA, and PC systems throughout this research paper (Johnson & Hsiao, 1992). By SA1 system, for example, we mean the SA1 receptors (the Merkel-neurite complex), the SA1 afferent nerve fiber population, and all the ascending and central neuronal pathways that convey the SA1 signal used for memory and perception. We do not mean to imply that there is no central convergence between these systems or that the systems do not overlap.
SA1 afferent fibers branch repeatedly before they lose their myelin and terminate in the basal layer of the epidermis. There, they are enveloped by specialized (Merkel) epidermal cells that enfold the unmyelinated ends of the SA1 axons (Iggo &Andres, 1982). Although there are synapse-like junctions between the Merkel cells and the axon terminals, action potentials appear to arise as the result of mechanosensitive ion channels in the bare nerve endings (Diamond, Mills, & Mearow, 1988; Ogawa, 1996). SA1 afferents innervate the skin of the fingerpad densely and have small receptive fields. As a consequence of these two properties, they transmit a high-resolution spatial neural image of a stimulus contacting the fingerpad. A striking response property of these afferents is surround suppression (Vega-Bermudez & Johnson, 1999b), which is similar to surround inhibition. In the central nervous system, surround inhibition in the receptive field of visual or somatosensory neurons is the result of an excitatory center and an inhibitory surround produced by inhibitory synaptic mechanisms. Surround inhibition makes a neuron sensitive to local curvature and—depending on the balance between excitation and inhibition—insensitive to uniform stimulation. In touch, surround suppression confers similar response properties to SA1 afferents; instead of being based on synaptic mechanisms, however, it is based entirely on mechanoreceptor sensitivity to a specific component of tissue strain near the nerve ending (strain energy density or a closely related component of strain; Phillips & Johnson, 1981; Srinivasan & Dandekar, 1996).As a consequence, SA1 afferents fire vigorously to points, edges, and curvature, and these responses are suppressed by the presence of stimuli in the surrounding skin (illustrated in Figure 8.2).Also, because of surround suppression, SA1 afferents are minimally responsive to uniform skin indentation; therefore, local spatial features such as edges and curves are represented strongly in the neural image conveyed by the peripheral SA1 population response. Combined psychophysical and neurophysiological experiments (reviewed later in this research paper) indicate that the SA1 system is responsible for form and texture perception.
RA afferent fibers also branch repeatedly as they near the epidermis. Each RA afferent ends in 30–80 Meissner’s corpuscles (Johnson,Yoshioka, &Vega-Bermudez, 2000), which occur in dermal pockets between the sweat duct and adhesive ridges (Guinard, Usson, Guillermet, & Saxod, 2000; Munger & Ide, 1988). This puts Meissner’s corpuscles, which lie in the dermis, as close to the surface of the epidermis as is possible (Quilliam, 1978); this may account, in part, for the RA’s greater sensitivity to minute skin deformation relative to SA1 afferents, whose receptors are located on the tips of the sweat-duct ridges. RA afferents innervate the skin of the fingerpad more densely than do the SA1 afferents. Based on combined data from humans and monkeys (Darian-Smith & Kenins, 1980; Johansson & Vallbo, 1976; Johansson & Vallbo, 1979b), which are not significantly different, the best estimates at the fingertip are 100 SA1 and 150 RA afferents/cm2. This greater innervation density suggests a role for RA afferents in the two-dimensional representation of stimulus form, but RA afferents resolve the spatial details of tactilestimulimorepoorlythandoSA1afferents.Theirreceptive field sizes depend strongly on stimulus intensity and are muchlargerthanSA1receptivefieldsatindentationlevelsthat occur in ordinary tactile experience. The striking feature of RA responses is their sensitivity to minute skin motion. The effective operating range of indentations for RAs is about 4–400 m; the comparable SA1 range is about 15–1,500 m or more (Blake, Johnson, & Hsiao, 1997; Johansson, 1979; Mountcastle, Talbot, & Kornhuber, 1966; Vega-Bermudez & Johnson, 1999a). Thus, the SA1 and RA response properties are complementary. The RA and SA1 systems are in some ways like the scotopic and photopic systems in vision. The RA system, like the scotopic system, has greater sensitivity but poorer spatial resolution and limited dynamic range. The SA 1 system, like the photopic system, is less sensitive but has higher spatial resolution and operates over a wider dynamic range. The neural response properties of RA afferents make them ideally suited for motion perception. In fact, combined psychophysical and neurophysiological studies show that the RA system is responsible for the perception of events that produce low-frequency, low-amplitude skin motion; that includes the detection of microscopic surface features, the detection of low-frequency vibration, and the detection of slip, which is critical for grip control (reviewed in Johnson et al., 2000).
Each PC afferent terminates in a single Pacinian corpuscle, which occurs in the dermis or the deeper tissues. The history, structure, and electrophysiological properties of this receptor are reviewed by Bell, Bolanowski, and Holmes (1994). The most striking feature of the PC response is its extreme sensitivity, which derives from mechanosensitive ion channels located in the unmyelinated ending within the corpuscle. Pacinian corpuscles respond with action potentials to vibratory amplitudes as small as 3 nm applied directly to the corpuscle (Bolanowski & Zwislocki, 1984) and 10 nm applied to the skin (Brisben, Hsiao, & Johnson, 1999). The corpuscles comprise multiple layers of fluid-filled sacs; these sacs act as a cascade of high-pass filters that shield the unmyelinated ending from the large, low-frequency deformations that accompany most manual tasks (Hubbard, 1958; Loewenstein & Skalak, 1966). If not for this intense filtering, the transducer, which is two orders of magnitude more sensitive than the other mechanoreceptive transducers, would be overwhelmed by most cutaneous stimuli. Because of their extreme sensitivity,receptivefieldboundariesaredifficulttodefine.SomePCs have receptive fields that encompass an entire hand or even an entire arm; other PC’s have receptive fields restricted to a single phalanx. There are about 2,500 Pacinian corpuscles in the human hand and they are about twice as numerous in the fingers as in the palm (about 350 per finger and 800 in the palm; reviewed in Brisben et al., 1999). Because of its small numbers and its large receptive fields, the PC population transmits little (if any) useful information about the spatial properties of a stimulus. Instead, it transmits very effectively information communicated by vibrations in objects, tools, or probes held in the hand (reviewed in Johnson et al., 2000).
SA2 afferents are distinguished from SA1 afferents by four properties: (a) Their receptive field areas are about five times larger than SA1 receptive fields and have borders that are not clearly demarcated (Johansson & Vallbo, 1980); (b) they are about six times less sensitive to skin indentation (Johansson & Vallbo, 1979a; Johansson, Vallbo, & Westling, 1980); (c) they are two to four times more sensitive to skin stretch (Edin, 1992); and (d) their interspike intervals are more uniform (Chambers, Andres, von Duering, & Iggo, 1972; Edin, 1992). SA2 afferents are thought to end in Ruffini complexes (Iggo & Andres, 1982), although the association of afferents with these response properties with a specific receptor is not as secure as it is with other cutaneous mechanoreceptors. Both SA1 and SA2 afferents respond to forces orthogonal and parallel to the skin surface, but between them the SA1 afferents are biased toward responsiveness to orthogonal forces, and SA2 afferents are biased toward parallel forces (Macefield, Hager-Ross, & Johansson, 1996). The poor SA2 responses to raised dot patterns (e.g., Braille patterns in Figure 8.4) and to curved surfaces suggest that they play no role in form perception (Phillips, Johansson, & Johnson, 1990). Because of SA2 responses to curved surfaces, Goodwin, Macefield, and Bisley (1997, p. 2887) conclude that “SA2 responses are unlikely to signal information to the brain about the local shape of an object.” Because of their sensitivity to skin stretch, SA2s are well suited to signaling lateral forces such as active forces pulling on an object held in the hand. An interesting possibility is that they send a neural image of the pattern of skin stretch to the central nervous system when the hand moves about and that this neural image plays a significant or possibly even the dominant role in our perception of hand conformation (reviewed in Johnson et al., 2000) and of the direction of motion of an object moving across the skin (Olausson, Wessberg, & Kakuda, 2000).
Form and Texture Perception
The distinction between form and texture perception is that form perception concerns perception of the geometric structure of a surface or object, whereas texture perception corresponds to the subjective feel of a surface or object. Form perception depends on the specific geometry of a surface or object; texture perception depends on its distributed, statistical properties. Form perception has many dimensions; texture perception has only two or possibly three dimensions (see this research paper’s section entitled “Texture Perception”). Form perception can be studied with objective methods (i.e., the subject’s responses can be scored for accuracy); texture perception cannot. A sheet of Braille text provides an example of the distinction. After scanning a Braille passage with the fingertip, a Braille reader can be asked to report the content of the passage or to report how rough it felt. The subject’s character- or word-recognition abilities can be scored for accuracy; the report of roughness cannot. Similarly, reports about the spacing or density of the Braille dots would be a product of form and not of texture perception.
Form Perception
Form perception is constant over a wide range of stimulus conditions. The ability to discriminate object or surface features and the capacity for pattern recognition at the fingertip are the same whether the object is contacted by active touch or is applied to the passive hand. Form perception is affected only marginally by whether the object is stationary or moving relative to the skin; it is unaffected by scanning speed up to 40mm/s; it is unaffected by contact force, at least over the range from0.2–1.0N; and it is affected only marginally by the height (relief) of spatial features over a wide range of heights (Johnson & Lamb, 1981; Loomis, 1981, 1985; Phillips, Johnson, & Browne, 1983; Vega-Bermudez, Johnson, & Hsiao,1991).The evidence presented in the following discussion shows that the SA1system is responsible for form perception.
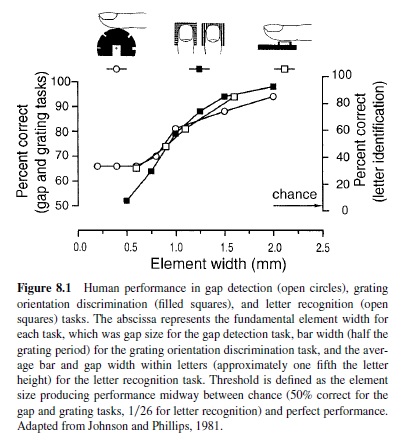
Figure 8.1 shows the results of three psychophysical studies of the human ability to discriminate stimuli with the distal pad of the index finger. In all three studies, the element width that resulted in performance midway between chance and perfect discrimination was between 0.9 and 1.0 mm, which is close to the theoretical limit set by the innervation density of SA1 and RAprimary afferents at the fingertip.Acuity declines progressively from the index finger to the fifth finger (Vega-Bermudez & Johnson, 2001) and it declines progressively with age (Sathian, Zangaladze, Green, Vitek, & DeLong, 1997; J. C. Stevens & Choo, 1996; Wohlert, 1996). Whether these declines are due to differences in innervation density is not known. Spatial acuity at the lip and tongue is significantly better than that at the fingertip (Essick, Chen, & Kelly, 1999; Sathian & Zangaladze, 1996; Van Boven & Johnson, 1994). Tactile spatial acuity is the same in human and nonhuman primates (Hsiao, O’Shaughnessy, & Johnson, 1993).
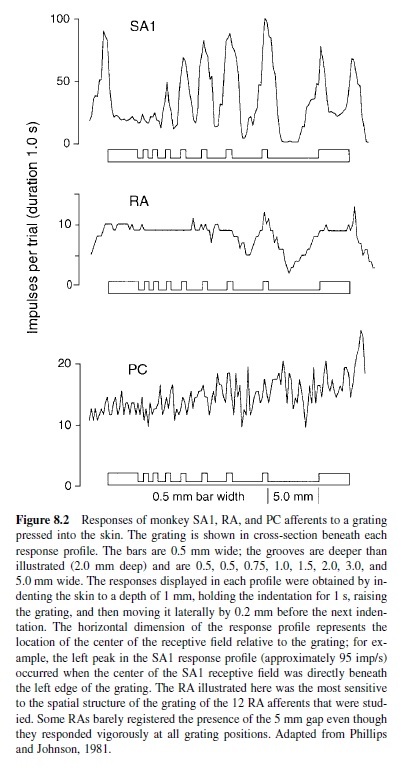
The ability to discriminate gratings and letters with element widths around 1 mm means that at least one of the afferent systems must sustain a neural image with 1 mm resolution or better. That requires an innervation density of at least one afferent per square mm; it also requires that individual afferents be able to resolve the spatial details with a resolution that accounts for human discrimination performance. Neither the PC system nor the SA2 system comes close on either score (Johansson & Vallbo, 1979b). Note that the human performance illustrated in Figure 8.1 begins to rise above chance at element sizes around 0.5 mm, which means that either the SA1 or RA system must begin to resolve spatial detail at about 0.5 mm. Evidence that only the SA1 afferents account for the human performance illustrated in Figure 8.1 comes from neurophysiological experiments in which SA1 and RA afferents were studied with periodic gratings. SA1 responses to a periodic grating convey information about spatial structure when the groove and ridge widths are 0.5 mm wide (Figure 8.2). When the grooves and ridges are 1 mm wide, SA1s provide a robust neural image of the stimulus. In contrast, RAs require grooves that are 3 mm wide or more before their responses begin to distinguish a grating from a flat surface; most RAs fail even to register grooves 3 mm wide. The RA response illustrated in Figure 8.2 was the most sensitive to spatial detail of the 12 RAs that were studied. PC afferents were unable to resolve grooves that were 5 mm wide (Figure 8.2). Kops and Gardner (1996) obtained nearly identical results with an Optacon, which is a dense array of vibrotactile probes designed as a reading aid for the blind (Bliss, 1969).
The human ability to recognize raised letters of the alphabet when the letter heights are near threshold is illustrated in Figure 8.3. In this figure, we also show the response of a typical SA1 afferent to repeated scans of the same raised letters (Vega-Bermudez et al., 1991). In that study, Vega-Bermudez et al. (1991) showed that there was no detectable difference in human performance between active and passive touch and that the confusion matrix shown in Figure 8.3 is characteristic of human letter recognition performance across a wide range of stimulus conditions. Even the detailed patterns of identifications and errors were identical between active and passive touch. Accuracies in letter recognition ranged from 15% (N) to 98% (I) and more than 50% of the confusions were confined to 7% of all possible confusion pairs (22 out of 325 possible confusion pairs), which are enclosed in boxes in Figure 8.3. The confusions in all but 5 of those 22 pairs are highly asymmetric ( p < 0.001). The frequency of occurrence of letters in English bears no relationship to the rates of correct responses, false positives, or total responses. Furthermore, excluding I, J, and L, which had high hit rates and low false-positive rates, there was no relationship between hit and false-positive rates. All of this evidence suggests that the response patterns are not the result of cognitive biases.

The pattern of confusions seems to be explained by the response of SA1 afferents to the letters, which is illustrated at the bottom of Figure 8.3. For example, B is identified as B on only 18% of trials; instead, it is called D 50% more often than it is called B. Conversely, D is virtually never called B (Figure 8.3 top). This pattern of confusion can be explained by the SA1 surround suppression mechanism discussed earlier, which suppresses the response to the central, horizontal bar of the B. The neural representation of the B resembles a D more than it does a B (see rasters at bottom of Figure 8.3), which accounts for the strong bias in the psychophysical studies towards the letter D. For another example, C is often called G or Q, but G and Q are almost never called C. An explanation is that the participant learns quickly that internal and trailing features are often represented weakly or not at all; therefore, when the participant is presented with the letter C, lack of the features expected of a G or a Q in the neural representation is not a strong reason to not respond G or Q. Conversely, the strong representation of the distinctive features of the G and Q make confusion with a C unlikely. The performance illustrated in Figure 8.3 is for naive subjects in their first testing session. Performance improves steadily on repeated testing (Vega-Bermudez et al., 1991). One possible reason for the improvement is that subjects may learn the idiosyncracies of the neural representations (e.g., as soon as a participant recognizes the distinctive feature of the G in the neural representation, he or she is less likely to mistake a C for a G).

Figure 8.4 shows the responses of typical human cutaneous afferent responses to Braille symbols (top row) scanned over their receptive fields (Phillips et al., 1990). The responses of human afferents to these raised-dot patterns are indistinguishable from the responses of monkey afferents to similar patterns (Johnson & Lamb, 1981). SA1 afferents provide a sharp, isomorphic representation of the Braille patterns, RA provide a less sharply defined isomorphic representation, and PCs and SA2’s provide no useful spatial information.
Neurophysiological studies with the Optacon show that it activates the RA and PC systems well but not the SA1 system; furthermore, these studies demonstrate that only the RAsystem can account for spatial pattern recognition performance with the Optacon (Gardner & Palmer, 1989; Palmer & Gardner, 1990). Therefore, the psychophysical studies employing the Optacon (J. C. Craig & Rollman, 1999) are studies of the form-processing capacity of the RA system. The results of those studies are exactly what one would predict based on the RA responses illustrated in Figure 8.2. For example, humans cannot discriminate the orientation of an Optacon grating pattern until the gaps in the grating exceed 5 mm width (Kops & Gardner, 1996).
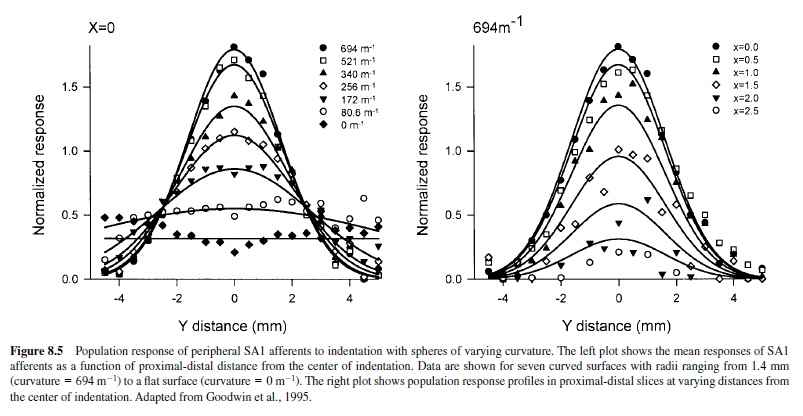
Studies of curvature perception also implicate the SA1 system in form processing (Goodwin, Browning, & Wheat, 1995; Goodwin, John, & Marceglia, 1991; Goodwin & Wheat, 1992a, 1992b). Those studies show that estimates of curvature are unaffected by changes in contact area and force, and—conversely—estimates of force are unaffected by changes in curvature. This latter finding is particularly surprising, considering that SA1 firing rates are strongly affected by curvature and that SA1 mean firing rates are the most likely neural code for the perception of force (Goodwin et al., 1995; LaMotte & Srinivasan, 1993; Srinivasan & LaMotte, 1987). The psychophysical observations showing that curvature perception is unaffected by changes in contact area or force suggest that the spatial profile of neural activity is used for the perception of curvature and that a different neural code (e.g., total discharge rate) is used for the perception of force. Only the SA1 population response provides a veridical representation of curvature that can account for the psychophysical observations (Dodson, Goodwin, Browning, & Gehring, 1998; Goodwin et al., 1995). The SA1 population responses to a wide range of curvatures are shown in Figure 8.5. RAs respond poorly to such stimuli and provide no signal that might account for the ability of humans to discriminate curvature (Goodwin et al., 1995; Khalsa, Friedman, Srinivasan, & LaMotte, 1998; LaMotte, Friedman, Lu, Khalsa, & Srinivasan, 1998).
Texture Perception
Multidimensional scaling studies have shown that texture perception includes soft-hard and smooth-rough as independent perceptual dimensions—surface hardness and roughness can occur in almost any combination, and they account for most or all of texture perception (Hollins, Bensmaïa, Karlof, & Young, 2000; Hollins, Faldowski, Rao, & Young, 1993). A third dimension (sticky-slippery) improves the multidimensional scaling fit in some subjects. Thus, it appears that texture perception has two strong dimensions and possibly a third weaker dimension.
Roughness. Roughness perception has been studied extensively (Blake, Hsiao, & Johnson, 1997; Connor, Hsiao, Phillips, & Johnson, 1990; Connor & Johnson, 1992; Hollins et al., 2000; Lederman, 1974; Meenes & Zigler, 1923; Meftah, Belingard, & Chapman, 2000; Sathian, Goodwin, John, & Darian-Smith, 1989; S. S. Stevens & Harris, 1962; Yoshioka, Dorsch, Hsiao, & Johnson, 2001). These studies demonstrate that roughness perception is unidimensional (the test of unidimensionality being the ability to assign numbers on a unidimensional continuum and to make greater-than and less-than judgments); that it depends on element height, diameter, shape, compliance, and density; and that the relationship between roughness and the different element parameters is complex and nonlinear. Important early observations were that scanning velocity, contact force, and friction between the finger and a surface have minor or no effects on roughness magnitude judgments (Lederman, 1974; Taylor & Lederman, 1975).
The neural mechanisms of roughness perception have been studied in a series of combined psychophysical and neurophysiological studies (reviewed in Johnson et al., 2000). These studies suggest strongly that the neural mechanisms underlying roughness perception depend on the spatial variation (mean absolute difference) in firing rates between SA1 afferents with receptive field centers separated by 2–3 mm. This conclusion was derived through the use of the method of successive falsification (Platt, 1964; Popper, 1959), whereby psychophysical data from human participants were tested against neurophysiological data recorded from monkeys using the exact same stimuli and stimulus conditions. In this method, multiple working hypotheses are proposed, and a hypothesis is rejected only when there is no consistent (oneto-one) relationship between the neural measure and human performance. The favored hypothesis is the one that survives the hypothesis testing.The first study (Connor et al., 1990) rejected neural codes based on mean firing rate; the second study (Connor & Johnson, 1992) rejected all neural codes that depend on the temporal fluctuations in firing rates of the afferent fibers; the third study (Blake, Hsiao, et al., 1997) rejected all neural codes based on the firing of RA afferent fibers; the fourth study (Yoshioka et al., 2001) rejected all neural codes based on the firing of PC afferent fibers. The only neural code that consistently accounts for human roughness perception over surfaces with individual elements spaced from 0.1 to 6.2 mm apart and heights from 0.28 to 2.0 mm is one based on spatial variation of firing among SA1 afferent fibers. The correlation between the psychophysical roughness magnitude estimates and spatial variation in the SA1 discharge was greater than 0.97 in all four studies (see Figure 8.6).
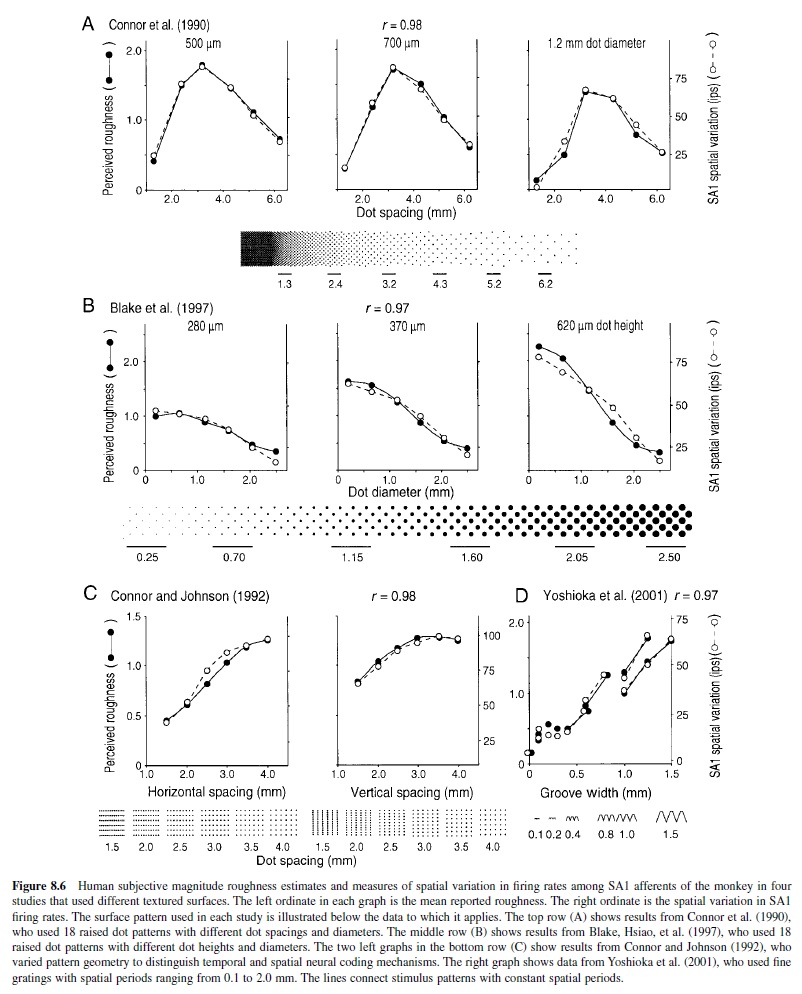
Recent studies, reviewed in the following discussion, have shown that a subpopulation of neurons in area 3b of primary somatosensory cortex (SI) have receptive fields composed of spatially separated regions of excitation and inhibition. These neurons have discharge rates that are proportional to the difference in discharge rates that one would expect between SA1 afferent fibers with receptive fields separated by 2–3 mm (DiCarlo & Johnson, 2000)—that is, the mechanism underlying their firing rates is exactly the one that was discussed earlier as the neural basis for roughness judgments. The mean firing rate of a population of such neurons would correspond closely to subjects’ roughness judgments; moreover, like roughness perception, the mean firing rate would be affected only secondarily by scanning velocity and contact force (DiCarlo & Johnson, 1999). These results suggest that roughness perception may be based on the responses of a subpopulation of neurons in area 3b.
Softness. Softness (or its reciprocal, hardness) is the second major dimension of texture (Hollins et al., 2000). Softness is the subjective impression of the progressive change in conformation of a surface to the contours of the fingers that accompanies changes in contact force. The perception of softness is not to be confused with the perception of compliance. Although both are dependent on changes in contact force, compliance is a physical property of the surface being touched—consequently, discrimination of compliance, like dot spacing, is objective (i.e., can be scored for accuracy). In contrast, softness, like roughness, is a subjective sense. Perceived softness does not depend on the relationship between force and object displacement—the fact that the space bar on a computer keyboard gives way easily (is compliant) does not make it soft. Asoft object conforms to the finger or hand as it is manipulated, but conformation is not sufficient; the keyboard keys that are molded to conform to the skin of a fingertip feel as hard as flat keys. A working hypothesis is that softness is signaled by the rate of growth of contact area with contact force and by the uniformity of pressure across the contact area.
The neural mechanisms of softness perception have not been studied systematically. Except for a study by Harper and Stevens (1964), most psychophysical studies have focused on the objective ability to discriminate compliance. Harper and Stevens showed that subjective softness judgments were related to the compliance of their test objects by a power function and that hardness and softness judgments were reciprocallyrelated.Themostextensivestudyoftheabilityofhumans to discriminate compliance is by Srinivasan and LaMotte (1995), who used cutaneous anesthesia and various modes of stimulus contact to show that cutaneous information alone is sufficient to discriminate the compliance of objects with deformable surfaces. Subjects discriminate softness when an object is applied to the passive, immobile finger as accurately as they do when they actively palpate the object. Moreover, the study showed that this ability is unaffected when the velocity and force of application are randomized.
There are no combined psychophysical and neurophysiological experiments that systematically address the neural mechanisms of softness perception, but the likely mechanism can be inferred from what we know about the response properties of each of the afferent types. Just as in roughness perception, the a priori possibilities are intensive, temporal, or spatial neural codes in one or more of the cutaneous afferent populations. Intensive codes are unlikely because random changes in velocity and force, which do not affect discrimination performance, have strong effects on afferent impulse rates (Srinivasan & LaMotte, 1996). Purely temporal codes seem unlikely because perceived softness (or hardness) is based on perceived changes in object form with changing contact force. Hence, the most likely mechanism is that softness is based on the dynamic changes in the profile of the population response of the SA1 afferents that occur when the finger comes into contact with a surface.
Texture Perception With a Probe. When we use a tool or probe, we perceive distant events almost as if our fingers were present at the working surfaces of the tool or probe. An early demonstration of this was made by Katz (1925/1989), who showed that we can discriminate one textured surface from another as well with a probe as with a finger applied directly to the surfaces. He showed further that this capacity is lost when vibrations in the probe are damped. A recent study (Brisben et al., 1999) has shown that when subjects grasp a probe, transmitted vibrations with amplitudes less than 10 nm at the hand can be detected (the mean is 30 nm; see Figure 8.7). Only the PC system can account for this capacity.
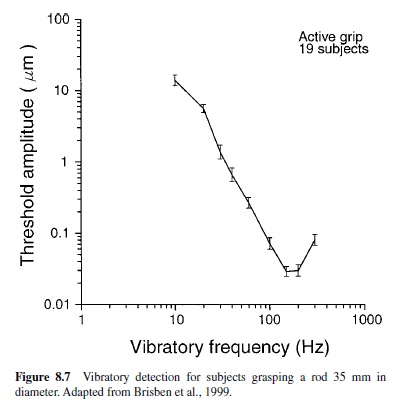
The hypothesis that the PC system is responsible for the perception of vibrations transmitted through an object held in the hand supposes not only that Pacinian receptors detect the transmitted vibration, but also that the PC population transmits a neural representation of the vibratory signal sufficient to account for this perceptual capacity. Work on the human ability to detect and discriminate complex vibratory stimuli supports this idea by showing that we are sensitive to the temporal structure of high-frequency stimuli that only activate PC afferents (Formby, Morgan, Forrest, & Raney, 1992; Lamore, Muijser, & Keemink, 1986; Weisenberger,1986);forexample, humans can discriminate the frequency with which a 250-Hz carrier stimulus is modulated for modulation frequencies as high as 60 Hz (Formby et al., 1992). In contrast, Bensmaïa and Hollins (2000) have shown that the discrimination of complex waveforms composed of high frequencies is poor; they have suggested that RA afferents may play a role in temporal coding.
Central Mechanisms of Form and Texture Perception
Central Pathways
A principle of organization within the somatosensory system is that neurons responsible for the different aspects of sensory perception are separated into separate anatomical pathways that are modality specific. The division begins at the level of the peripheral nerves and continues as axons leaving the dorsal root ganglion send their projections into the spinal cord. Axons entering the spinal cord separate into two parallel paths (Figure 8.8). One—the dorsal-column-medial-lemniscal pathway—contains large- and medium-diameter myelinated fibers that ascend in the ipsilateral dorsal column to synapse on neurons in the dorsal column nuclei (DCN). This pathway is responsible for conveying information concerning both mechanoreceptive and proprioceptive function. The other pathway—the spinothalamic tract (STT)—conveys information from unmyelinated C fibers and small myelinated A-delta fibers that enter and terminate in the dorsal horn of the spinal cord. From there, the axons of the STTcross the midline to ascend in the anterolateral quadrant of the spinal cord. This pathway is primarily responsible for conveying information about pain and temperature. The division of function continues as second-order neurons in the DCN and dorsal horn send their axons to the thalamic nuclei (A. D. Craig & Dostrovsky, 1999; Jones, 1990; Jones & Friedman, 1982; Perl, 1998; Poggio&Mountcastle, 1960). For the dorsal-column-mediallemniscal pathway, the second-order neurons terminate at the contralateral ventral posterior lateral nucleus of the thalamus (VPL), where afferents from the deep tissues are segregated from the cutaneous inputs (Jones & Friedman, 1982). Neurons in VPLthen send their projections to primary (SI) somatosensory cortex, whereas neurons in ventral posterior inferior nucleus (VPI) project to secondary (SII) somatosensory cortex (Friedman, Murray, O’Neill, & Mishkin, 1986; Jones & Burton, 1976; Jones & Friedman, 1982).
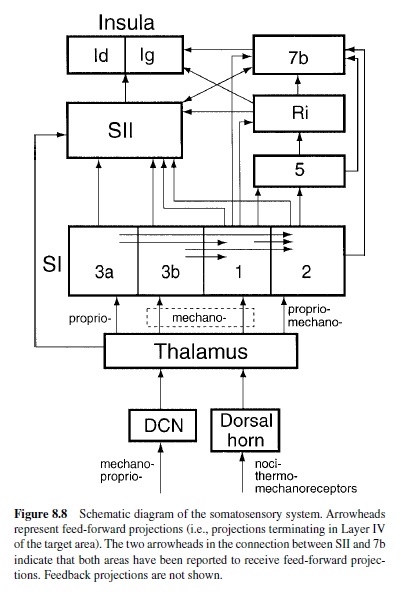
SI cortex is composed of four cytoarchitectonic areas (3a, 3b, 1, and 2) that receive the majority of their inputs from VPL. The connectivity is highly specific, with neurons in 3a and 2 receiving inputs from neurons in VPL that respond to proprioceptive stimuli and neurons in 3b and 1 receiving inputs from neurons in VPL that respond to cutaneous stimuli. The four areas of SI are also interconnected; neurons in 3a send a large projection to area 2 and a small projection to 1, neurons in 3b send their outputs to 1 and 2, and 1 sends its outputs to 2 (for a review, see Felleman & Van Essen, 1991). Based on these connections alone, one can infer that areas 3b and 1 are important for mechanoreceptive functions and that neurons in 3a and 2 are important for functions like stereognosis that require proprioceptive information (Landgren, Silfvenius, & Olsson, 1984).
There are two main projections from SI. One is toward areas 5, Ri, and 7b; the other is toward SII. SII also receives inputs from Ri and 7b and along with these areas sends its outputs to two separate divisions of the insula. Although in Figure 8.8 we have depicted SII as a single area, it is composed of two or more areas (reviewed later in this research paper). A working hypothesis with extensive support from both neurophysiological and ablation studies is that the main pathway underlying form and texture perception proceeds from the thalamus to SI cortex, then to SII cortex, and finally to the insula (Schneider, Friedman, & Mishkin, 1993).
SI Cortex
The segregation of function between the SA1, RA, SA2, and PC systems suggests that these afferent systems remain segregated to some degree in the central nervous system. However, considering how distinctly separate are the functions of the four afferent systems, it is surprising that there is little evidence to support the notion that these systems are segregated within SI cortex. One of the few definite claims that can be made is that 3b is important for form perception. In the remainder of this section on SI cortex, we review the neural mechanisms of form processing in area 3b.
Area 3b. Removal of area 3b produces profound behavioral deficits in all somatosensory tasks tested, whereas removal of other SI areas appears to produce more specific deficits in the tactile discrimination of textures (area 1) and three-dimensional forms (area 2; Randolph & Semmes, 1974). Neurophysiological studies have shown that 3b has smaller receptive fields (Paul, Goodman, & Merzenich, 1972; Sur, Garraghty, & Bruce, 1985; Sur, Merzenich, & Kaas, 1980) and a higher proportion of cells responding to static skin indentation (Paul et al., 1972; Sur, Wall, & Kaas, 1984) than do other SI cortical areas. However, none of these observations is especially indicative of 3b playing arole in spatial information processing. The smaller receptive fields in 3b are almost certainly a consequence of 3b being at an earlier stage of processing within the somatosensory pathways (Bankman, Johnson, & Hsiao, 1990; Johnson, Hsiao, & Twombly, 1995).Acontinued response to sustained, steady indentation does imply that neurons in 3b receive inputs from slowly adapting afferents, but the converse is not true—a transient response may result from lack of slowly adapting input or, just as likely, delayed inhibition that shuts off the response to a sustained input (Andersson, 1965; DiCarlo & Johnson, 2000; Gardner & Costanzo, 1980; Innocenti & Manzoni, 1972; Sur et al., 1984).
Aclue that 3b is specialized for processing spatial information comes from the relative expansions of the representations of the digits in areas 3b and 1. The cortical magnification factors (unit cortical area per unit body surface area) in areas 3b and 1 are approximately equal over most of the postcentral gyrus, except in the finger region, where spatial acuity is highest and the magnification in 3b climbs to approximately five timesthatinarea1(Suretal.,1980).Anothercluecomesfrom the response properties of neurons in 3b. When neurons with receptive fields on the fingerpads are stimulated with scanned, complex spatial stimuli, almost all of them yield responses that are more complex than can be accounted for by simple, excitatory receptive fields. It is evident from these responses that many neurons in 3b are responding to specific spatial features of the stimuli (Bankman et al., 1990; Hsiao, Johnson, Twombly, & DiCarlo, 1996; Johnson et al., 1995; Phillips, Johnson, & Hsiao, 1988). Responses of two SA neurons in area 3b are illustrated in Figure 8.9. The neuron illustrated in the top two rows of Figure 8.9 is clearly responding to the orientations of letter segments. The neuron illustrated in the bottom two rows has a complex response that is not easily interpreted; for example, the central response to the letters appears to be completely suppressed by the horizontal bars within the B, E, and F but not the H. Thus, the neuronal responses in 3b suggest specialization for form processing rather than (for example) motion processing.

Receptive Fields in Area 3b. A recent series of studies with controlled, scanned stimuli has confirmed that there is little or no directional selectivity in area 3b and that 3b neuronal discharge rates are affected only mildly by changes in stimulus velocity across the skin (DiCarlo & Johnson, 1999, 2000; DiCarlo, Johnson, & Hsiao, 1998). They also show, however, that all neurons in 3b are selectively responsive to particular spatial patterns of stimulation, that they are sensitive to the orientation of these patterns, and that this selectivity is shaped as much by inhibition as it is by excitation. Previous studies have generally failed to identify this inhibition using a simple probe because it is manifested only as a reduction of the response to a stimulus that simultaneously contacts both the excitatory and inhibitory parts of the receptive field. Ninety-five percent of 3b neuronal receptive fields have three components: (a) a single, central excitatory region of short duration (10 ms at most), (b) one or more inhibitory regions that are adjacent to and synchronous with the excitation, and (c) a larger inhibitory region that overlaps the excitation partially or totally and is delayed with respect to the first two components (by 30 ms on average). The remaining 5% have two or more regions of excitation.
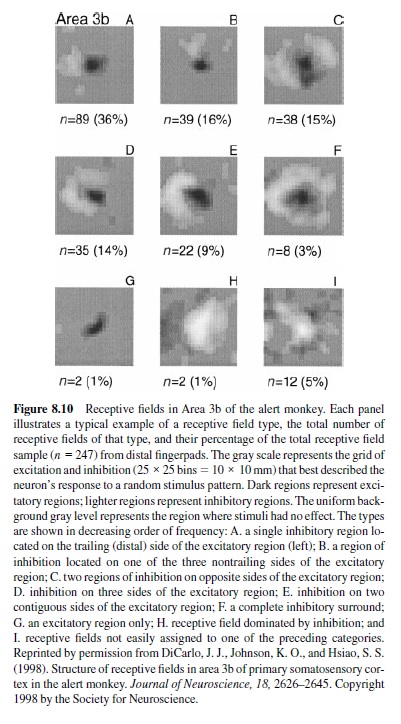
The receptive fields of 247 area 3b neurons mapped with scanned, random-dot stimuli are illustrated in Figure 8.10, which shows that virtually all receptive fields are characterized by a single central region of excitation with inhibition on one, two, or three sides. Surround inhibition occurred rarely. The inhibitory area was, on average, about 30% larger than the excitatory area (means were 18 and 14 mm2) and—like the excitatory area—varied greatly (from 1–47 mm2). The inhibitory mass (absolute value of inhibition integrated over the entire inhibitory field), like the excitatory mass (comparable definition), varied by 50 to 1 between neurons (125–6,830 mass units; mean 1,620 mass units). There was no evidence of clustering into distinct receptive field types. The distributions of excitatory and inhibitory areas and masses were all Gaussian in logarithmic coordinates (i.e., lognormal); the excitatory and inhibitory masses were more closely correlated ( = 0.56) than were the areas ( = 0.26). Receptive fields mapped in this way accurately predict neuronal responses to stimulus features such as orientation (DiCarlo & Johnson, 2000).
Delayed Inhibition. Area 3b neurons have two striking response properties that are not evident in Figure 8.10. The first is that the spatiotemporal structure of their neuronal responses and the spatial structures of their receptive fields are virtually unaffected by the velocity with which a stimulus moves across the skin or (conversely) how rapidly a finger is scanned over a surface for velocities up to at least 80 mm/s (DiCarlo & Johnson, 1999)—that is, the spatial structure of responses like the ones illustrated in Figure 8.9 are unaffected by changes in scanning velocity. Increasing scanning velocity causes a marked increase in the intensities of the excitatory and inhibitory subfields without affecting their geometries; this results in increased firing rates without any loss of the response selectivity conferred by the receptive field geometry. The mechanism of this increased responsiveness with increased velocity lies in an interaction between the excitation and the delayed inhibition (DiCarlo & Johnson, 1999).
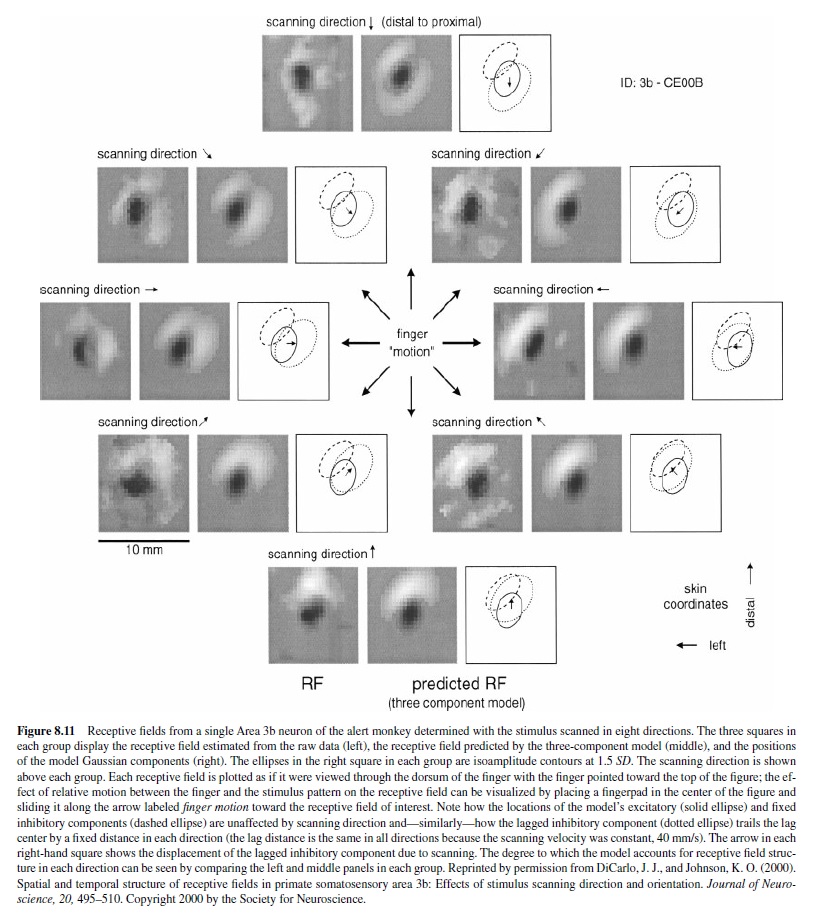
The delayed inhibition confers a second property, which causes the geometry of the receptive field to be strongly scanning-direction-dependent (DiCarlo & Johnson, 2000). A typical example of a response that is dependent on scanning direction is shown in Figure 8.11. The receptive field at the left of each group of three receptive field diagrams is the receptive field obtained directly from the neuron’s responses to random-dot stimuli scanned in one of the eight directions. This figure illustrates that regardless of scanning direction, there is a fixed region of inhibition distal and left of the central region of excitation. It can also be seen that there is a region of inhibition displaced in the scanning direction (opposite to the finger motion) from the region of excitation. To visualize this more clearly, each neuron’s response was fitted with a three-component receptive field model comprising a Gaussian excitatory region and two Gaussian inhibitory regions—one to simulate the region of fixed, synchronous inhibition and one to simulate the region of delayed inhibition. This model is illustrated by the simulated receptive field in the central panel in each group of three panels and by the corresponding diagram in the right panel. The degree to which the model description accounts for the observed receptive fields can be seen by comparing the model receptive field in the central panel and the actual receptive field in the left panel. This comparison shows that the three-component model explains the effect of scanning direction on receptive field shape well. The correlation between the model and observed receptive fields averaged 0.81 in 62 neurons studied with four or more scanning directions. Neurons with lower correlations all had a third, fixed region of inhibition not accounted for by the three-component model (i.e., they would have been described by the three components enumerated previously if the model had allowed for more than one region of fixed inhibition).
Functional Implications. The wide range of receptive field geometries and the wide range of responses to complex, scanned stimuli found in 3b shows that the initial, isomorphic neural representation of spatial form that prevails in the periphery gives way to an altered form of representation in which neuronal responses represent the presence of specific features. The more complex responses observed in SII cortex (Hsiao et al., 1996) suggest that 3b is an intermediate step in a series of transformations leading to a more complex form of representation (Bankman et al., 1990; DiCarlo et al., 1998; Johnson et al., 1995).
The fixed inhibitory components of each neuron’s receptive field interacts with the central excitation to act as a spatial filter, conferring selectivity for particular spatial features or patterns regardless of scanning direction and velocity. For example, when the fixed inhibition lies on two adjacent sides, the neuron is more responsive to corners that protrude into the excitatory subfield without activating the inhibitory subregions. When the fixed inhibitory subfield occupies a single location on one side of the excitatory subfield, both tend to be elongated and to lie parallel to one another; as a result, the neuron is more responsive to edges oriented parallel to the two subregions (DiCarlo & Johnson, 2000; DiCarlo et al., 1998).
The delayed inhibitory component serves three functions. First, it confers sensitivity to stimulus gradients in the scanning direction, regardless of that direction. The delayed inhibition suppresses the response to uniform surfaces and thereby emphasizes the effects of spatial or temporal novelty. When scanning the finger over a surface, features first activate the regions of excitation and fixed inhibition and then 30 ms later activate the lagged inhibition. Second, when the delayed inhibition is centered on the excitation it produces a progressiveincreaseindischargeratewithincreasingscanningvelocity. The acquisition of tactile spatial information by scanning one’s finger over a surface compensates for the very limited field of view provided by a single fingerpad. It is clearly an advantage to be able to scan one’s fingers over an object or a surface rapidly without loss of information. Psychophysical experiments demonstrate that performance in pattern recognition is unaffected as scanning rate increases from 20 to 40 mm/s, and then only a small loss is observed as the rate is increased to 80 mm/s (Vega-Bermudez et al., 1991). In the absence of a compensatory mechanism, rapid scanning has a substantialcost.Asscanningvelocityincreases,eachstimulus element spends less time within the receptive field (reduced dwell time) and the element is represented by fewer action potentials. The delayed inhibition provides a compensatory mechanism that increases the firing rate with increasing scanning velocity. As velocity increases, the delayed inhibition lags progressively to expose more excitation. Consequently, the excitatory and inhibitory components of the receptive field grow rapidly in intensity, with no effect on receptive field geometry. The result is a representation of spatial form that is invariant with scanning velocity and that is more intense than it would be without this mechanism (DiCarlo & Johnson, 1999).Third—andleastsignificant—isthatdirectionsensitivity occurs when the delayed inhibition is displaced from the center of excitation (Barlow & Levick, 1965; DiCarlo & Johnson, 2000; Gardner & Costanzo, 1980; Warren, Hämäläinen, & Gardner, 1986); this is because motion in the direction of the displacement exposes progressively more excitation, which produces a progressively greater discharge rate. Motion in the opposite direction shifts the delayed inhibition over the center of excitation, thereby reducing the discharge rate. The center of the delayed inhibition in area 3b is, with few exceptions, close to the center of excitation, which may explain why so few neurons in 3b exhibit directional selectivity. When the center of the delayed inhibition is displaced from the center of excitation, it predicts the neuron’s directional selectivity accurately (DiCarlo & Johnson, 2000).
SII Cortex
SII was first described in the early 1940s. Adrian (1941) reported a second tactile area in the ectosylvian gyrus of the cat; he hypothesized that this area was specialized for processing information from the animal’s paw. Woolsey (1943) studied this area more extensively and reported that it also existed in both dogs and monkeys; furthermore, he found that it contained a complete somatotopically organized representation of the body surface.Since those initial studies, the functional role and detailed organization of SII has remained elusive. Neurophysiological mapping studies (Robinson & Burton, 1980c; Whitsel, Petrucelli, & Werner, 1969), anatomical tracer studies in macaque monkeys (Burton, Fabri, & Alloway, 1995; Friedman & Murray, 1986; Krubitzer, Clarey, Tweedale, Elston, & Calford, 1995), and recent neural imaging studies in humans (Disbrow, Roberts, & Krubitzer, 2000) show that SII cortex extends approximately 10 mm across the upper bank of the lateral sulcus and that what was initially identified as SII cortex comprises two separate areas—SII and parietal ventral cortex (Krubitzer et al., 1995), which are also called SIIp and SIIr (Burton & Sinclair, 1996; Whitsel et al., 1969).Although these areas have separate somatotopic maps, the functional roles that they play in tactile perception are not known.
Evidence from anatomical tracer studies (Friedman & Murray, 1986) and from studies in which areas of cortex were selectively deactivated by cooling suggest that SI and SII process information in parallel (G. M. Murray, Zhang, Kaye, Sinnadurai, Campbell, & Rowe, 1992; Zhang et al., 1996), whereas lesion (Pons, Garraghty, Friedman, & Mishkin, 1987) and neurophysiological studies (Burton & Sinclair, 1990; Hsiao et al., 1993) suggest otherwise. In the cooling studies, Rowe and his colleagues were unable to abolish SII responses by cooling SI cortex, which suggests that SI and SII process information in parallel (Zhang et al., 1996). In contrast, Pons et al. (1987) provided strong evidence for serial processing. When SI is ablated, neurons in SII become silent; when SII is ablated, neurons in SI remain active (Pons, Wall, Garraghty, Cusick, & Kaas, 1987). In other studies they showed that selective lesions in the subregions that constitute SI result in modality-selective deficits in the response properties of neurons in SII (Garraghty, Pons, & Kaas, 1990; Pons, Garraghty, & Mishkin, 1992). Neurophysiological evidence from single-cell studies supports the hypothesis that SII lies at a processing stage higher than that of SI. Neurons in SI tend to have small, simple receptive fields confined to one (area 3b) or a few digits on a single hand (Iwamura, Tanaka, Sakamoto, & Hikosaka, 1983; Sur et al., 1984). In contrast, neurons in SII tend to have larger and more complex receptive fields that span multiple digits on one or both hands (reviewed in the next section of this research paper). A third line of evidence supporting the serial hypothesis is that almost all of the neurons in SII are strongly affected by the animal’s focus of attention, suggesting that this area is closely related to higher cognitive aspects of tactile perception.
Role of SII in Tactile Processing. SII cortex plays an important role in texture and two- and three-dimensional shape processing. Ettlinger and his colleagues demonstrated that monkeys with ablations of SII were unable to perform almost all tactile tasks that required touch (Garcha & Ettlinger, 1978; Ridley & Ettlinger, 1976). In addition, they found that animals that had SII cortex ablated were impaired in their ability to learn new tasks and were unable to do tasks that required intermanual transfer of information. Murray and Mishkin (1984) showed that when SII was ablated, monkeys were unable to discriminate texture (hard vs. soft and rough vs. smooth), form (square vs. diamond and convex vs. concave shapes), and object orientation (horizontal vs. vertical) and were significantly impaired in size discrimination (small vs. large objects). Furthermore, they showed that SII and not area 5 is important for these functions because animals with lesions confined to area 5 showed no deficits on the same tasks (Murray & Mishkin, 1984).
Neurophysiological studies support the idea that SII is further along the pathways responsible for processing tactile form. Neurons in SII tend to have large receptive fields that often span multiple digits on the same hand (receptive fields < 10 cm2, Robinson & Burton, 1980a) and respond to stimulation of both hands (Burton & Carlson, 1986; Cusick, Wall, Felleman, & Kaas, 1989; Robinson & Burton, 1980a; Whitsel et al., 1969; Sinclair & Burton, 1993). Stimulus selectivity varies widely in SII cortex. Some SII neurons are activated by light touch, whereas others require complex stimuli (Burton & Sinclair, 1990, 1991; Chapman, Zompa, Williams, Shenasa, & Jiang, 1996; Ferrington & Rowe, 1980; Fitzgerald, Lane, Yoshioka, Nakama, & Hsiao, 1999; Hsiao et al., 1993; Pruett, Sinclair, & Burton, 2000; Robinson & Burton, 1980b; Sinclair & Burton, 1993).
Recently Fitzgerald et al. (1999) used oriented bars as stimuli and found that approximately 30% of the neurons in SII cortex showed orientation-tuned responses. The preferred orientations of these neurons were approximately uniformly distributed across the eight orientations that were tested. In addition, they found that most of those neurons showed similar orientation tuning preferences between phalanges and between digits. They hypothesized that these orientationsensitive neurons could be part of a network that is involved in generating a positionally invariant representation of tactile stimuli or could be involved in representing shape information for objects that span multiple digits. The results from studies in SII using embossed letters suggest that SII lies at a processing stage higher than that of SI. First, unlike neurons in SI, none of the neurons in SII show isomorphic responses. Instead, the responses tend to be highly nonisomorphic and feature selective. Examples of two neuronal responses to scanned letters are shown in Figure 8.12. These neurons responded well to trailing features and to specific local features such as the leading parts of the letters C and D but not to the leading parts of the letters B and E (Figure 8.12).

Several studies have investigated the responses of neurons in SII to vibration and to textured patterns. Two separate studies that used vibratory stimuli report that many neurons in SII show phase-locking to both low- and high-frequency vibrations (Burton & Sinclair, 1991; Ferrington & Rowe, 1980). The degree of phase-locking was greater for SII neurons than for SI neurons, suggesting that vibratory information may bypass SI cortex (Ferrington & Rowe, 1980). Studies using active and passive scanning of gratings suggest that neurons in SII are sensitive to textured stimuli (Jiang, Tremblay, & Chapman, 1997; Pruett et al., 2000; Sinclair & Burton, 1993). These studies demonstrated that the firing rates of many neurons in SII show monotonic increases (or decreases) in firing rate as the spatial period of a grating is increased. One of these studies (Jiang et al., 1997) reported that the majority of the neurons in SII that responded to the textured surfaces showed responses that were more related to the differences in spatial periods than to the spatial periods themselves.
Attention
Psychophysics of Tactile Attention
Selective attention plays an important role in sensory processing. Cross-modal studies in which attention is switched between touch and a visual or auditory stimulus suggest that attention has a limited capacity. In one series of studies, Chapman and her colleagues asked participants to direct or divide their focus of attention between the onset of either a 40-Hz vibration or a light (Post & Chapman, 1991) or detect the occurrence of a textured stimulus and a light (Zompa & Chapman, 1995). In both studies, the stimuli were preceded with cues that directed participants’attention either toward or away from the correct stimulus or directed their attention to both stimuli (divided attention task). They found that the attentional resources are limited and that subjects had the shortest reaction times when cued correctly, intermediate reaction times when given the neutral cue, and the longest reaction times when cued incorrectly. Different results are found when attention is switched between tactile stimuli at multiple skin sites. Shiffrin, Craig, and Cohen (1973) had participants detect the occurrence of vibrations presented at three different locations on the body (thenar eminence of the right hand, tip of left index finger, and the forearm); participants were cued either to direct their attention to the correct site or to divide their attention between the three sites. In contrast to the cross-modal studies, they found that there was no difference in performance in the directed and divided conditions. Similarly, J. C. Craig (1985) found that there were minimal differences in performance when attention was directed to a single hand or was divided between the two hands. These studies suggest that under certain circumstances the capacity of attention may be large.
Focus of Attention
Several studies show that attention may function like a cognitive spotlight, so to speak. Lakatos and Shepard (1997) reported that the time it takes to discriminate the presence or absence of a tactile stimulus depends on the distance between the cued site and the test locations. A surprising finding was that the critical distance was not related to the somatotopic distance (i.e., anatomical distance along the body surface between the tested sites) but instead was related to the straightline distance between the two test sites (Lakatos & Shepard, 1997). Driver and Grossenbacher (1996) reported similar results in studies in which they delivered vibrotactile stimuli to hands that were either together or spread apart. They also showed that reaction times for detecting stimuli on the relevant hand decreased when participants oriented their heads toward the hand; these results were unaffected when the participants were blindfolded. These studies indicate that the coordinate system for attention is based on an internal representation of extrapersonal space and that proprioception plays a role in selective attention.
The spotlight of attention appears to have a minimum aperture size. The fundamental finding is that vibrotactile stimuli presented on a single hand tend to interact. For example, J. C. Craig (1985) showed that there is a decrement in performance when individuals are told to attend to a single finger and competing stimuli are presented on other fingers of the same hand; if identical stimuli are presented on target and nontarget fingers, then performance is improved. Similar results were reported by Franzén, Markowitz, and Swets (1970), who showed that vibrotactile stimuli presented to two fingers on the same hand interact. These studies suggest that the hand may be under a single attentional focus. In contrast, there appears to be minimal interference when stimuli are presented simultaneously to both hands. Evans and Craig (1991) reported that nontarget distractors have minimal effects when stimuli are presented to fingers on different hands. Similarly, J. C. Craig (1985) asked participants to combine pattern fragments presented to two fingers on the same or opposite hands. He found that individuals integrated the pattern fragments with greater accuracy when they were presented to two fingers on opposite hands than when they were presented to adjacent fingers of the same hand. He also showed that the participants’ performance on tasks in which the stimuli were presented on fingers on opposite hands was unaffected by the spatial separation between the two hands.
Using a search task, Klatzky, Lederman, and O’Neil (1996) showed that features based on surface properties (e.g., rough-smooth, hard-soft, and cool-warm) and surface discontinuities tended to have flat reaction time slopes (RT slopes < 30 ms per distractor item), whereas other features such as bar orientation have steep reaction time slopes. Features based on three-dimensional contours had intermediate reaction times. Whang, Burton, and Shulman (1991) and Sathian and Burton (1991) performed similar studies using vibratory and textured stimuli; they showed that selective attention is only minimally required to detect a change in texture or vibration (Sathian & Burton, 1991; Whang et al., 1991). Recently Sinclair, Kuo, and Burton (2000) showed that attention can selectively separate vibration frequency and roughness from stimulus duration.
Effects of Attention on Responses of Neurons in the Somatosensory System
The focus of attention has been shown to affect even the earliest stages of information processing within the somatosensory system. Hayes, Dubner, and Hoffman (1981) reported that the neuronal responses in the dorsal horn of the spinal cord were modulated by the attentional focus of animals performing thermal and light discrimination tasks. Although Poranen and Hyvärinen (1982) reported no effects of attention on neurons in the ventral posterior area of the thalamus (VP), other studies have reported that attention has small but significant effects on the response of these neurons. Bushnell, Duncan, Dubner, Fang, and He (1984) found a small number of neurons that were affected by the animal’s attentional focus, and recently Morrow and Casey (2000) showed that a small but significant number of neurons (7/18) in VPL showed attention-modulated responses.
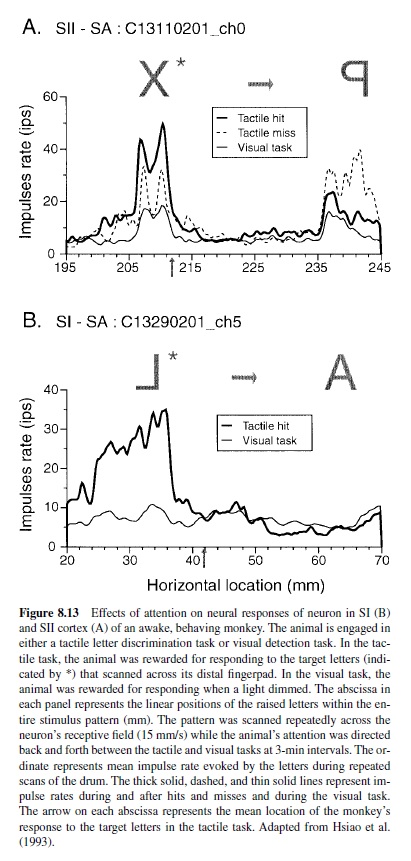
Similar descriptions of attention-modulated responses have been reported in SI cortex. In an early study, Hyvärinen, Poranen, and Jokinen (1980) recorded from neurons in SI cortex of animals trained to perform a vibration detection task; they found that about 8% of the neurons in area 3b and about 22% of the neurons in area 1 were affected by the animals’ focus of attention. Furthermore, they reported that all of these neurons showed enhanced responses. In another study, Poranen and Hyvärinen (1982) found that the attentional state had minimal effects on the neural responses. In both of those studies, the effects could have been due to arousal because the effects were assessed by comparing neural responses when the animals were performing the detection task with responses while the animal sat passively. Hsiao et al. (1993) controlled for arousal effects by recording from animals trained to perform a tactile letter discrimination task and a visual detection task. During both tasks, the same tactile stimuli were scanned across the distal pads of the animals’ hands, and the effects of attention were assessed by comparing the responses recorded during the two attentional states. They found that 50% of SI neurons had increased firing rates when the animal attended to the tactile stimulus. Furthermore, the firing rates were increased only during the presentation of letters that counted toward receiving a reward. For example, during the reward period or during timeout periods triggered by a false-positive response, the neural response rates were the same as they were when the animal was performing the visual task. Figure 8.13 shows examples of the effect of attention on the responses of neurons in SI and SII cortex. This figure shows the responses of neurons to a target letter (*) and the following nontarget letter that the animal learned was behaviorally irrelevant (i.e., the animal learned it could not receive a reward for letters following target letters). Both neurons showed a significant increase in neural activity when the animal performed the tactile task relative to the visual task. The effect was not simply a change in gain; it was also a change in the form of the neural response to the stimulus (Figure 8.13). Burton and Sinclair (2000) recorded from neurons in animals trained to detect a change in vibratory amplitude. They also reported that about 50% of neurons in SI cortex are affected by the animal’s focus of attention.
Recent studies indicate that neurons in SI cortex have altered responses during the delay period in a memory-related task (Zhou & Fuster, 1996, 1997). In those studies, animals were trained to perform either a tactile-tactile match or a cross-modal visual-tactile match. They found that during the delay period between the stimuli, many neurons in all four areas of SI cortex showed sustained rate changes. These studies suggest that neurons in SI may participate in the short-term memory of tactile stimuli.
Studies in SII show that attention has more profound effects on the responses of neurons in SII than in SI cortex. Based on multiunit recordings, Poranen and Hyvärinen (1982) found that whereas attention had minimal effects on the responses of neurons in SI, all of the neurons in SII were affected. Similarly, Hsiao et al. (1993) reported that 80% of the neurons in SII were affected by attention and that the effects were divided between increased (58%) and decreased (22%) neural firing rates. Burton, Sinclair, Hong, Pruett, and Whang (1997) studied animals performing a vibration detection task and found that 45% of the neurons in SII cortex were affected by the animal’s focus of attention. They showed that the responses were generally suppressed during the early phases and enhanced during the late phases of a trial.
A modification of impulse rate is only one of the ways that attention might affect the responses of a neuron. Studies in SI and SII cortex employing multiple electrodes have shown that synchronous firing between neurons may play an important role in tactile information processing (A. Roy, Steinmetz, Hsiao, Johnson, & Niebur, 2001; S. Roy & Alloway, 1999; Steinmetz et al., 2000). S. Roy and Alloway (1999) observed that more than 60% of the neuron pairs in SI fired synchronously and that the degree of synchronous firing increased when stimuli were moved across the skin. Steinmetz et al. (2000) investigated the synchrony of neuronal discharge in SII cortex in animals trained to switch their attention between a visual task and a tactile discrimination task. They reported that a large fraction of the neuron pairs in SII cortex fired synchronously and—more important—that the degree of synchrony was affected by the animal’s focus of attention (Figure 8.14). On average 17% of the neuron pairs that fired synchronously showed changes in the degree of synchrony when the animal switched its focus of attention from the visual to the tactile task. In addition, the percentage of neurons that showed changes in synchrony differed between the three animals in a predictable way. Thirty-five percent of the neuron pairs showed changes in synchrony in the animal performing the most difficult tactile task, whereas only 9% of the neuron pairs were affected in the animal performing the easiest task.
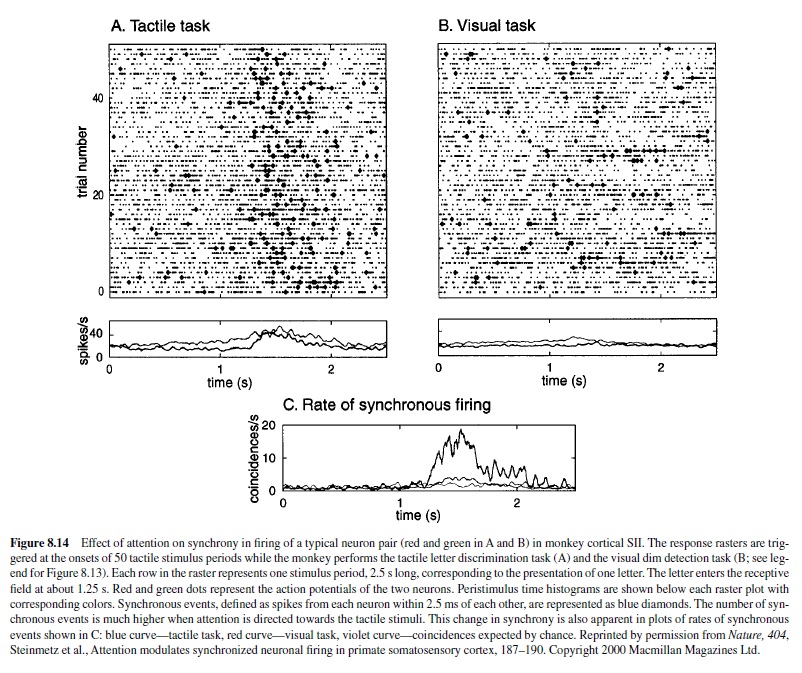
These findings suggest that attention may operate by changing the synchrony of firing of selected neurons. Synchronous firing between neurons that have a common target produces larger excitatory postsynaptic potentials in the target neuron than does asynchronous firing and is therefore more effective at driving the target neuron. Hence, when the firing in a subpopulation is made more synchronous by attention, the message contained in that subpopulation is selected preferentially for further processing. Changes in firing synchrony may be the neural correlate of selective attention.
Summary
In this research paper we have reviewed the peripheral neural mechanisms of tactile sensation, the neural mechanisms in SI and SII cortex, and the effects of attention on information processing in the somatosensory pathways. Three decades of combined psychophysical and neurophysiological experiments suggest a sharp division of function among the four cutaneous afferent systems that innervate the human hand. The SA1 system provides a high-quality neural image of the spatial structure of objects and surfaces that contact the skin; this is the basis of form and texture perception. The RAsystem provides a neural image of motion signals from the whole hand. From this image, the brain extracts information that is critical for grip control and also information about the motion of objects contacting the skin.The PC system provides a neural image of vibrations transmitted from objects grasped in the hand. The SA2 system provides a neural image of skin stretch, which changes as hand conformation changes. The evidence is less secure, but the most likely hypothesis is that the brain extracts information from the SA2 population response about hand conformation and the direction of motion of objects moving across the skin. SI and SII cortex transform information from the four afferent systems to an altered form of representation that is still not well understood.
This review concentrated on the processing of spatially patterned stimuli. Neurons in Area 3b of SI cortex, which is the first cortical somatosensory processing area, have small receptive fields restricted to a single finger. These neurons have responses that are selective for the local spatial features of a stimulus. Neurons in SII cortex have more complex responses and often have receptive fields that span multiple fingers or both hands. Neurons in SII cortex are—like neurons in SI cortex—orientation selective, and the orientation selectivity is consistent over multiple fingers on the same hand. These responses may represent a mechanism that confers position invariance (i.e., responses to a stimulus are the same regardless of position), or they may indicate a mechanism that integrates information across multiple fingers, which is necessary for object recognition. Selective attention is an important mechanism in sensory processing. Neurophysiological studies show that attention affects neuronal responses at the very first level of processing within the somatosensory pathways (the dorsal horn of the spinal cord) and becomes progressively more important at higher levels within the system. In SII cortex, the focus of attention has a profound affect on the responses of single neurons and on the temporal patterning of neural activity between neurons.
Bibliography:
- Adrian, E. D. (1941). Afferent discharges to the cerebral cortex from peripheral sense organs. Journal of Physiology, 100, 159–191.
- Andersson, S. A. (1965). Intracellular postsynaptic potentials in the somatosensory cortex of the cat. Nature, 205, 297–298.
- Bankman, I. N., Johnson, K. O., & Hsiao, S. S. (1990). Neural image transformation in the somatosensory system of the monkey: Comparison of neurophysiological observations with responses in a neural network model. Cold Spring Harbor Symposia on Quantitative Biology, 55, 611–620.
- Barlow, H. B., & Levick, W. R. (1965). The mechanism of directionally selective units in rabbit’s retina. Journal of Physiology, 178, 477–504.
- Bell, J., Bolanowski, S. J., & Holmes, M. H. (1994). The structure and function of Pacinian corpuscles: A review. Progress in Neurobiology, 42, 79–128.
- Bensmaïa, S. J., & Hollins, M. (2000). Complex tactile waveform discrimination. Journal of the Acoustical Society of America, 108, 1236–1245.
- Blake, D. T., Hsiao, S. S., & Johnson, K. O. (1997). Neural coding mechanisms in tactile pattern recognition: The relative contributions of slowly and rapidly adapting mechanoreceptors to perceived roughness. Journal of Neuroscience, 17, 7480–7489.
- Blake, D. T., Johnson, K. O., & Hsiao, S. S. (1997). Monkey cutaneous SAI and RA responses to raised and depressed scanned patterns: Effects of width, height, orientation, and a raised surround. Journal of Neurophysiology, 78, 2503–2517.
- Bliss, J. C. (1969). A relatively high-resolution reading aid for the blind. IEEE Transactions on Man-Machine Systems, 10, 1–9.
- Bolanowski, S. J., & Zwislocki, J. J. (1984). Intensity and frequency characteristics of Pacinian corpuscles: Pt. 1. Action potentials. Journal of Neurophysiology, 51, 793–811.
- Brisben, A. J., Hsiao, S. S., & Johnson, K. O. (1999). Detection of vibration transmitted through an object grasped in the hand. Journal of Neurophysiology, 81, 1548–1558.
- Burton, H., & Carlson, M. (1986). Second somatic sensory cortical area (SII) in a prosimian primate, Galago crassicaudatus. Journal of Comparative Neurology, 247, 200–220.
- Burton, H., Fabri, M., & Alloway, K. D. (1995). Cortical areas within the lateral sulcus connected to cutaneous representations in areas 3b and 1: A revised interpretation of the second somatosensory area in macaque monkeys. Journal of Comparative Neurology, 355, 539–562.
- Burton, H., & Sinclair, R. J. (1990). Second somatosensory cortical area in macaque monkeys: I. Neuronal responses to controlled, punctate indentions of glabrous skin on the hand. Brain Research, 520, 262–271.
- Burton, H., & Sinclair, R. J. (1991). Second somatosensory cortical area in macaque monkeys: II. Neuronal responses to punctate vibrotactile stimulation of glabrous skin on the hand. Brain Research, 538, 127–135.
- Burton, H., & Sinclair, R. J. (1996). Somatosensory cortex and tactile perceptions. In L. Kruger (Ed.), Pain and touch (pp. 105– 177). San Diego, CA: Academic Press.
- Burton, H., & Sinclair, R. J. (2000). Tactile-spatial and cross-modal attention effects in the primary somatosensory cortical areas 3b and 1-2 of rhesus monkeys. Somatosensory and Motor Research, 17, 213–228.
- Burton, H., Sinclair, R. J., Hong, S. Y., Pruett, J. R., & Whang, K. C. (1997). Tactile-spatial and cross-modal attention effects in the second somatosensory and 7b cortical areas of rhesus monkeys. Somatosensory and Motor Research, 14, 237– 267.
- Bushnell, M. C., Duncan, G. H., Dubner, R., Fang, L., & He, L. F. (1984). Activity of trigeminothalamic neurons in medullary dorsal horn of awake monkeys trained in a thermal discrimination task. Journal of Neurophysiology, 52, 170–187.
- Chambers, M. R., Andres, K. H., von Duering, M., & Iggo, A. (1972). The structure and function of the slowly adapting type II mechanoreceptor in hairy skin. Quarterly Journal of Experimental Physiology, 57, 417–445.
- Chapman, C. E., Zompa, I. C., Williams, S. R., Shenasa, J., & Jiang, W. (1996). Factors influencing the perception of tactile stimuli during movement. In O. Franzén, R. S. Johansson, & L. Terenius (Eds.), Somesthesis and the neurobiology of the somatosensory cortex (pp. 307–320). Basel, Switzerland: Birkhäuser Verlag.
- Connor, C. E., Hsiao, S. S., Phillips, J. R., & Johnson, K. O. (1990). Tactile roughness: neural codes that account for psychophysical magnitude estimates. Journal of Neuroscience, 10, 3823–3836.
- Connor, C. E., & Johnson, K. O. (1992). Neural coding of tactile texture: comparisons of spatial and temporal mechanisms for roughness perception. Journal of Neuroscience, 12, 3414– 3426.
- Craig, A. D., & Dostrovsky, J. O. (1999). Medulla to thalamus. In P. D. Wall & R. Melzack (Eds.), Textbook of pain (pp. 183–214). Hong Kong: Harcourt.
- Craig, J. C. (1985). Attending to two fingers: two hands are better than one. Perception and Psychophysics, 38, 496–511.
- Craig, J. C., & Rollman, G. B. (1999). Somesthesis. Annual Review of Psychology, 50, 305–331.
- Cusick, C. G., Wall, J. T., Felleman, D. J., & Kaas, J. H. (1989). Somatotopic organization of the lateral sulcus of owl monkeys: Area 3b, S-II, and a ventral somatosensory area. Journal of Comparative Neurology, 282, 169–190.
- Darian-Smith, I., & Kenins, P. (1980). Innervation density of mechanoreceptive fibers supplying glabrous skin of the monkey’s index finger. Journal of Physiology, 309, 147–155.
- Diamond, J., Mills, L. R., & Mearow, K. M. (1988). Evidence that the Merkel cell is not the transducer in the mechanosensory Merkel cell-neurite complex. Progress in Brain Research, 74, 51–56.
- DiCarlo, J. J., & Johnson, K. O. (1999). Velocity invariance of receptive field structure in somatosensory cortical area 3b of the alert monkey. Journal of Neuroscience, 19, 401–419.
- DiCarlo, J. J., & Johnson, K. O. (2000). Spatial and temporal structure of receptive fields in primate somatosensory area 3b: Effects of stimulus scanning direction and orientation. Journal of Neuroscience, 20, 495–510.
- DiCarlo, J. J., Johnson, K. O., & Hsiao, S. S. (1998). Structure of receptive fields in area 3b of primary somatosensory cortex in the alert monkey. Journal of Neuroscience, 18, 2626–2645.
- Disbrow, E., Roberts, T., & Krubitzer, L. (2000). Somatotopic organization of cortical fields in the lateral sulcus of Homo sapiens: evidence for SII and PV. Journal of Comparative Neurology, 418, 1–21.
- Dodson, M. J., Goodwin, A. W., Browning, A. S., & Gehring, H. M. (1998). Peripheral neural mechanisms determining the orientation of cylinders grasped by the digits. Journal of Neuroscience, 18, 521–530.
- Driver, J., & Grossenbacher, P. G. (1996). Multimodal spatial constraints on tactile selective attention. In T. Inui & J. McClelland (Eds.), Attention and performance: Vol. 16. Information integration in perception and communication (attention and performance). Cambridge, MA: MIT Press, 209–235.
- Edin, B. B. (1992). Quantitative analysis of static strain sensitivity in human mechanoreceptors from hairy skin. Journal of Neurophysiology, 67, 1105–1113.
- Essick, G. K., Chen, C. C., & Kelly, D. G. (1999). A letterrecognition task to assess lingual tactile acuity. Journal of Oral and Maxillofacial Surgery, 57, 1324–1330.
- Evans, P. M., & Craig, J. C. (1991). Tactile attention and the perception of moving tactile stimuli. Perception and Psychophysics, 49, 355–364.
- Felleman, D. J., & Van Essen, D. C. (1991). Distributed hierarchical processing in the primate cerebral cortex. Cerebral Cortex, 1, 1–47.
- Ferrington, D. G., & Rowe, M. J. (1980). Differential contributions to coding of cutaneous vibratory information by cortical somatosensory areas I and II. Journal of Neurophysiology, 43, 310–331.
- Fitzgerald, P. J., Lane, J.W.,Yoshioka,T., Nakama,T., & Hsiao, S. S. (1999). Multidigit receptive field structures and orientation tuning properties of neurons in SII cortex of the awake monkey [Abstract]. Society for Neuroscience Abstracts, 25,
- Formby, C., Morgan, L. N., Forrest, T. G., & Raney, J. J. (1992). The role of frequency selectivity in measures of auditory and vibrotactile temporal resolution. Journal of the Acoustical Society of America, 91, 293–305.
- Franzén, O., Markowitz, J., & Swets, J. A. (1970). Spatially-limited attention to vibrotactile stimulation. Perception and Psychophysics, 7, 193–196.
- Friedman, D. P., & Murray, E. A. (1986). Thalamic connectivity of the second somatosensory area and neighboring somatosensory fields of the lateral sulcus of the macaque. Journal of Comparative Neurology, 252, 348–373.
- Friedman, D. P., Murray, E. A., O’Neill, J. B., & Mishkin, M. (1986). Cortical connections of the somatosensory fields of the lateral sulcus of macaques: Evidence for a corticolimbic pathway for touch. Journal of Comparative Neurology, 252, 323–347.
- Garcha, H. S., & Ettlinger, G. (1978). The effects of unilateral or bilateral removals of the second somatosensory cortex (area SII): A profound tactile disorder in monkeys. Cortex, 14, 319–326.
- Gardner, E. P., & Costanzo, R. M. (1980). Temporal integration of multiple-point stimuli in primary somatosensory cortical receptive fields of alert monkeys. Journal of Neurophysiology, 43, 444–468.
- Gardner, E. P., & Palmer, C. I. (1989). Simulation of motion on the skin: I. Receptive fields and temporal frequency coding by cutaneous mechanoreceptors of Optacon pulses delivered to the hand. Journal of Neurophysiology, 62, 1410–1436.
- Garraghty, P. E., Pons, T. P., & Kaas, J. H. (1990). Ablations of areas 3b (SI proper) and 3a of somatosensory cortex in marmosets deactivate the second and parietal ventral somatosensory areas. Somatosensory and Motor Research, 7, 125–135.
- Goodwin, A. W., Browning, A. S., & Wheat, H. E. (1995). Representation of curved surfaces in responses of mechanoreceptive afferent fibers innervating the monkey’s fingerpad. Journal of Neuroscience, 15, 798–810.
- Goodwin, A. W., John, K. T., & Marceglia, A. H. (1991). Tactile discrimination of curvature by humans using only cutaneous information from the fingerpads. Experimental Brain Research, 86, 663–672.
- Goodwin, A. W., Macefield, V. G., & Bisley, J. W. (1997). Encoding of object curvature by tactile afferents from human fingers. Journal of Neurophysiology, 78, 2881–2888.
- Goodwin, A. W., & Wheat, H. E. (1992a). Human tactile discrimination of curvature when contact area with the skin remains constant. Experimental Brain Research, 88, 447–450.
- Goodwin, A. W., & Wheat, H. E. (1992b). Magnitude estimation of force when objects with different shapes are applied passively to the fingerpad. Somatosensory and Motor Research, 9, 339– 344.
- Guinard, D., Usson, Y., Guillermet, C., & Saxod, R. (2000). PS-100 and NF 70-200 double immunolabeling for human digital skin Meissner corpuscle 3D imaging. Journal of Histochemistry and Cytochemistry, 48, 295–302.
- Harper, R., & Stevens, S. S. (1964). Subjective hardness of compliant materials. Quarterly Journal of Experimental Psychology, 16, 204–215.
- Hayes, R. L., Dubner, R., & Hoffman, D. S. (1981). Neuronal activity in medullary dorsal horn of awake monkeys trained in a thermal discrimination task: II. Behavioral modulation of responses to thermal and mechanical stimuli. Journal of Neurophysiology, 46, 428–443.
- Hollins, M., Bensmaïa, S. J., Karlof, K., & Young, F. (2000). Individual differences in perceptual space for tactile textures: Evidence from multidimensional scaling. Perception and Psychophysics, 62, 1534–1544.
- Hollins, M., Faldowski, R., Rao, S., & Young, F. (1993). Perceptual dimensions of tactile surface texture: A multidimensionalscaling analysis. Perception and Psychophysics, 54, 697–705.
- Hsiao, S. S., Johnson, K. O., Twombly, I. A., & DiCarlo, J. J. (1996). Form processing and attention effects in the somatosensory system. In O. Franzén, R. S. Johansson, & L. Terenius (Eds.), Somesthesis and the neurobiology of the somatosensory cortex (pp. 229–247). Basel, Switzerland: Birkhäuser Verlag.
- Hsiao, S. S., O’Shaughnessy, D. M., & Johnson, K. O. (1993). Effects of selective attention of spatial form processing in monkey primary and secondary somatosensory cortex. Journal of Neurophysiology, 70, 444–447.
- Hubbard, S. J. (1958). A study of rapid mechanical events in a mechanoreceptor. Journal of Physiology, 141, 198–218.
- Hyvärinen, J., Poranen, A., & Jokinen, Y. (1980). Influence of attentive behavior on neuronal responses to vibration in primary somatosensory cortex of the monkey. Journal of Neurophysiology, 43, 870–882.
- Iggo, A., & Andres, K. H. (1982). Morphology of cutaneous receptors. Annual Review of Neuroscience, 5, 1–31.
- Innocenti, G. M., & Manzoni, T. (1972). Response patterns of somatosensory cortical neurones to peripheral stimuli: An intracellular study. Archives Italiennes de Biologie, 110, 322–347.
- Iwamura, Y., Tanaka, M., Sakamoto, M., & Hikosaka, O. (1983). Functional subdivisions representing different finger regions in area 3 of the first somatosensory cortex of the conscious monkey. Experimental Brain Research, 51, 315–326.
- Jiang, W., Tremblay, F., & Chapman, C. E. (1997). Neuronal encoding of texture changes in the primary and the secondary somatosensory cortical areas of monkeys during passive texture discrimination. Journal of Neurophysiology, 77, 1656– 1662.
- Johansson, R. S. (1979). Tactile afferent units with small and well demarcated receptive fields in the glabrous skin area of the human hand. In D. R. Kenshalo (Ed.), Sensory function of the skin in humans (pp. 129–145). New York: Plenum.
- Johansson, R. S., & Vallbo, Å. B. (1976). Skin mechanoreceptors in the human hand: An inference of some population properties. In Y. Zotterman (Ed.), Sensory functions of the skin in primates (pp. 171–184). Oxford, England: Pergamon Press.
- Johansson, R. S., & Vallbo, Å. B. (1979a). Detection of tactile stimuli: Thresholds of afferent units related to psychophysical thresholds in the human hand. Journal of Physiology, 297, 405–422.
- Johansson, R. S., & Vallbo, Å. B. (1979b). Tactile sensibility in the human hand: Relative and absolute densities of four types of mechanoreceptive units in glabrous skin. Journal of Physiology, 286, 283–300.
- Johansson, R. S., & Vallbo, Å. B. (1980). Spatial properties of the population of mechanoreceptive units in the glabrous skin of the human hand. Brain Research, 184, 353–366.
- Johansson, R. S., Vallbo, Å. B., & Westling, G. (1980). Thresholds of mechanosensitive afferents in the human hand as measured with von Frey hairs. Brain Research, 184, 343–351.
- Johnson, K. O., & Hsiao, S. S. (1992). Neural mechanisms of tactual form and texture perception. Annual Review of Neuroscience, 15, 227–250.
- Johnson, K. O., Hsiao, S. S., & Twombly, I. A. (1995). Neural mechanisms of tactile form recognition. In M. S. Gazzaniga (Ed.), The cognitive neurosciences (pp. 235–268). Cambridge, MA: MIT Press.
- Johnson, K. O., & Lamb, G. D. (1981). Neural mechanisms of spatial tactile discrimination: Neural patterns evoked by Braillelike dot patterns in the monkey. Journal of Physiology, 310, 117–144.
- Johnson, K. O., & Phillips, J. R. (1981). Tactile spatial resolution: I. Two-point discrimination, gap detection, grating resolution, and letter recognition. Journal of Neurophysiology, 46, 1177–1191.
- Johnson, K. O., Yoshioka, T., & Vega-Bermudez, F. (2000). Tactile functions of mechanoreceptive afferents innervating the hand. Journal of Clinical Neurophysiology, 17, 539–558.
- Jones, E. G. (1990). The thalamus. New York: Plenum.
- Jones, E. G., & Burton, H. (1976). Areal differences in the laminar distribution of thalamic afferents in cortical fields of the insular, parietal and temporal reigons of primates. Journal of Comparative Neurology, 168, 197–248.
- Jones, E. G., & Friedman, D. P. (1982). Projection pattern of functional components of thalamic ventrobasal complex upon monkey somatic sensory cortex. Journal of Neurophysiology, 48, 521–544.
- Katz, D. (1989). The world of touch (L. E. Krueger, Trans.). Hillsdale, NJ: Erlbaum. (Original work published 1925)
- Khalsa, P. S., Friedman, R. M., Srinivasan, M. A., & LaMotte, R. H. (1998). Encoding of shape and orientation of objects indented into the monkey fingerpad by populations of slowly and rapidly adapting mechanoreceptors. Journal of Neurophysiology, 79, 3238–3251.
- Klatzky, R. L., Lederman, S. J., & O’Neil, C. O. (1996). Haptic object processing: I. Early perceptual features. In O. Franzén, R. S. Johansson, & L. Terenius (Eds.), Somesthesis and the neurobiology of the somatosensory cortex (pp. 147–152). Basel, Switzerland: Birkhäuser Verlag.
- Kops, C. E., & Gardner, E. P. (1996). Discrimination of simulated texture patterns on the human hand. Journal of Neurophysiology, 76, 1145–1165.
- Krubitzer, L. A., Clarey, J., Tweedale, R., Elston, G., & Calford, M. B. (1995). A redefinition of somatosensory areas in the lateral sulcus of macaque monekeys. Journal of Neuroscience, 15, 3821–3839.
- Lakatos, S., & Shepard, R. N. (1997). Time-distance relations in shifting attention between locations on one’s body. Perception and Psychophysics, 59, 557–566.
- Lamore, P. J. J., Muijser, H., & Keemink, C. J. (1986). Envelope detection of amplitude-modulated high-frequency sinusoidal signals by skin mechanoreceptors. Journal of the Acoustical Society of America, 79, 1082–1085.
- LaMotte, R. H., Friedman, R. M., Lu, C., Khalsa, P. S., & Srinivasan, M. A. (1998). Raised object on a planar surface stroked across the fingerpad: Responses of cutaneous mechanoreceptors to shape and orientation. Journal of Neurophysiology, 80, 2446– 2466.
- LaMotte, R. H., & Srinivasan, M. A. (1993). Responses of cutaneous mechanoreceptors to the shape of objects applied to the primate fingerpad. Acta Psychologica, 84, 41–51.
- Landgren, S., Silfvenius, H., & Olsson, K. A. (1984). The sensorimotor integration in area 3a of the cat. Experimental Brain Research, 9(Suppl.), 359–375.
- Lederman, S. J. (1974). Tactile roughness of grooved surfaces: The touching process and the effects of macro- and microsurface structure. Perception and Psychophysics, 16, 385–395.
- Loewenstein, W. R., & Skalak, R. (1966). Mechanical transmission in a Pacinian corpuscle: An analysis and a theory. Journal of Physiology, 182, 346–378.
- Loomis, J. M. (1981). On the tangibility of letters and Braille. Perception and Psychophysics, 29, 37–46.
- Loomis, J. M. (1985). Tactile recognition of raised characters: A parametric study. Bulletin of the Psychonomic Society, 23, 18–20.
- Macefield, V. G., Hager-Ross, C., & Johansson, R. S. (1996). Control of grip force during restraint of an object held between finger and thumb: Responses of cutaneous afferents from the digits. Experimental Brain Research, 108, 155–171.
- Meenes, M., & Zigler, M. J. (1923). An experimental study of the perceptions of roughness and smoothness. American Journal of Psychology, 34, 542–549.
- Meftah, E. M., Belingard, L., & Chapman, C. E. (2000). Relative effects of the spatial and temporal characteristics of scanned surfaces on human perception of tactile roughness using passive touch. Experimental Brain Research, 132, 351–361.
- Morrow, T. J., & Casey, K. L. (2000). Attention-related, crossmodality modulation of somatosensory neurons in primate ventrobasal (VB) thalamus. Somatosensory and Motor Research, 17, 133–144.
- Mountcastle, V. B., Talbot, W. H., & Kornhuber, H. H. (1966). The neural transformation of mechanical stimuli delivered to the monkey’s hand. In A. V. de Reuck & J. Knight (Eds.), Touch, heat, pain and itch (pp. 325–351). London: Churchill.
- Munger, B. L., & Ide, C. (1988). The structure and function of cutaneous sensory receptors. Archives of Histology and Cytology, 51, 1–34.
- Murray, E. A., & Mishkin, M. (1984). Relative contributions of SII and area 5 to tactile discrimination in monkeys. Behavioural Brain Research, 11, 67–85.
- Murray, G. M., Zhang, H. Q., Kaye, A. N., Sinnadurai, T., Campbell, D. H., & Rowe, M. J. (1992). Parallel processing in rabbit first (SI) and second (SII) somatosensory cortical areas: Effects of reversible inactivation by cooling of SI on responses in SII. Journal of Neurophysiology, 68, 703–710.
- Ogawa, H. (1996). The Merkel cell as a possible mechanoreceptor cell. Progress in Neurobiology, 49, 317–334.
- Olausson, H., Wessberg, J., & Kakuda, N. (2000). Tactile directional sensibility: peripheral neural mechanisms in man. Brain Research, 866, 178–187.
- Palmer, C. I., & Gardner, E. P. (1990). Simulation of motion on the skin: IV. Responses of Pacinian corpuscle afferents innervating the primate hand to stripe patterns on the OPTACON. Journal of Neurophysiology, 64, 236–247.
- Paul, R. L., Goodman, H., & Merzenich, M. M. (1972). Alterations in mechanoreceptor input to Brodmann’s areas 1 and 3 of the postcentral hand area of Macaca mulatta after nerve section and regeneration. Brain Research, 39, 1–19.
- Perl, E. R. (1998). Getting a line on pain: is it mediated by dedicated pathways? Nature Neuroscience, 1, 177–178.
- Phillips, J. R., Johansson, R. S., & Johnson, K. O. (1990). Representation of Braille characters in human nerve fibers. Experimental Brain Research, 81, 589–592.
- Phillips, J. R., & Johnson, K. O. (1981). Tactile spatial resolution: II. Neural representation of bars, edges, and gratings in monkey primary afferents. Journal of Neurophysiology, 46, 1192–1203.
- Phillips, J. R., Johnson, K. O., & Browne, H. M. (1983). A comparison of visual and two modes of tactual letter resolution. Perception and Psychophysics, 34, 243–249.
- Phillips, J. R., Johnson, K. O., & Hsiao, S. S. (1988). Spatial pattern representation and transformation in monkey somatosensory cortex. Proceedings of the National Academy of Sciences, USA, 85, 1317–1321.
- Platt, J. R. (1964). Strong inference. Science, 146, 347–353.
- Poggio, G. F., & Mountcastle, V. B. (1960). Astudy of the functional contributions of the lemniscal and spinothalamic systems to somatic sensibility. Bulletin of the Johns Hopkins Hospital, 106, 266–316.
- Pons, T. P., Garraghty, P. E., Friedman, D. P., & Mishkin, M. (1987). Physiological evidence for serial processing in somatosensory cortex. Science, 237, 417–420.
- Pons, T. P., Garraghty, P. E., & Mishkin, M. (1992). Serial and parallel processing of tactual information in somatosensory cortex of rhesus monkeys. Journal of Neurophysiology, 68, 518–527.
- Pons, T. P., Wall, J. T., Garraghty, P. E., Cusick, C. G., & Kaas, J. H. (1987). Consistent features of the representation of the hand in area 3b of macaque monkeys. Somatosensory Research, 4, 309–331.
- Popper, K. (1959). The logic of scientific discovery. New York: Basic Books.
- Poranen, A., & Hyvärinen, J. (1982). Effects of attention on multiunit responses to vibration in the somatosensory regions of the monkey’s brain. Electroencephalography and Clinical Neurophysiology, 53, 525–537.
- Post, L. J., & Chapman, C. E. (1991). The effects of cross-modal manipulations of attention on the detection of vibrotactile stimuli in humans. Somatosensory and Motor Research, 8(2), 149–157.
- Pruett, J. R., Sinclair, R. J., & Burton, H. (2000). Response patterns in second somatosensory cortex (SII) of awake monkeys to passively applied tactile gratings. Journal of Neurophysiology, 84, 780–797.
- Quilliam, T. A. (1978). The structure of finger print skin. In G. Gordon (Ed.), Active touch (pp. 1–18). Oxford, UK: Pergamon Press.
- Randolph, M., & Semmes, J. (1974). Behavioral consequences of selective ablations in the postcentral gyrus of Macaca mulatta. Brain Research, 70, 55–70.
- Ridley, R. M., & Ettlinger, G. (1976). Impaired tactile learning and retention after removal of the second somatic sensory cortex (SII) in the monkey. Brain Research, 109, 656–660.
- Robinson, C. J., & Burton, H. (1980a). Organization of somatosensory receptive fields in cortical areas 7b, retroinsula, postauditory and granular insula of fascicularis. Journal of Comparative Neurology, 192, 69–92.
- Robinson, C. J., & Burton, H. (1980b). Somatic submodality distribution within the second somatosensory (SII), 7b, retroinsular, postauditory, and granular insular cortical areas of fascicularis. Journal of Comparative Neurology, 192, 93–108.
- Robinson, C. J., & Burton, H. (1980c). Somatotopographic organization in the second somatosensory area of fascicularis. Journal of Comparative Neurology, 192, 43–67.
- Roy, A., Steinmetz, P. N., Hsiao, S. S., Johnson, K. O., & Niebur, E. (2001). Synchrony: A neural correlate of somatosensory attention. Manuscript submitted for publication.
- Roy, S., & Alloway, K. D. (1999). Synchronization of local neural networks in the somatosensory cortex: A comparison of stationary and moving stimuli. Journal of Neurophysiology, 81, 999– 1013.
- Sathian, K., & Burton, H. (1991). The role of spatially selective attention in the tactile perception of texture. Perception and Psychophysics, 50, 237–248.
- Sathian, K., Goodwin, A. W., John, K. T., & Darian-Smith, I. (1989). Perceived roughness of a grating: Correlation with responses of mechanoreceptive afferents innervating the monkey’s fingerpad. Journal of Neuroscience, 9, 1273–1279.
- Sathian, K., & Zangaladze, A. (1996). Tactile spatial acuity at the human fingertip and lip: Bilateral symmetry and interdigit variability. Neurology, 46, 1464–1466.
- Sathian, K., Zangaladze, A., Green, J., Vitek, J. L., & DeLong, M. R. (1997). Tactile spatial acuity and roughness discrimination: Impairments due to aging and Parkinson’s disease. Neurology, 49, 168–177.
- Schneider, R. J., Friedman, D. P., & Mishkin, M. (1993). A modality-specific somatosensory area within the insula of the rhesus monkey. Brain Research, 621, 116–120.
- Shiffrin, R. M., Craig, J. C., & Cohen, E. (1973). On the degree of attention and capacity limitation in tactile processing. Perception and Psychophysics, 13, 328–336.
- Sinclair, R. J., & Burton, H. (1993). Neuronal activity in the second somatosensory cortex of monkeys (macaca mulatta) during active touch of gratings. Journal of Neurophysiology, 70, 331–350.
- Sinclair, R. J., Kuo, J. J., & Burton, H. (2000). Effects on discrimination performance of selective attention to tactile features. Somatosensory and Motor Research, 17, 145–157.
- Srinivasan, M. A., & Dandekar, K. (1996). An investigation of the mechanics of tactile sense using two-dimensional models of the primate fingertip. Journal of Biomechanical Engineering, 118, 48–55.
- Srinivasan, M. A., & LaMotte, R. H. (1987). Tactile discrimination of shape: Responses of slowly and rapidly adapting mechanoreceptive afferents to a step indented into the monkey fingerpad. Journal of Neuroscience, 7, 1682–1697.
- Srinivasan, M. A., & LaMotte, R. H. (1995). Tactual discrimination of softness. Journal of Neurophysiology, 73, 88–101.
- Srinivasan, M. A., & LaMotte, R. H. (1996). Tactual discrimination of softness: Abilities and mechanisms. In O. Franzén, R. S. Johansson, & L. Terenius (Eds.), Somesthesis and the neurobiology of the somatosensory cortex (pp. 123–135). Basel, Switzerland: Birkhäuser Verlag.
- Steinmetz, P. N., Roy, A., Fitzgerald, P. J., Hsiao, S. S., Johnson, K. O., & Niebur, E. (2000). Attention modulates synchronized neuronal firing in primate somatosensory cortex. Nature, 404, 187–190.
- Stevens, J. C., & Choo, K. K. (1996). Spatial acuity of the body surface over the life span. Somatosensory and Motor Research, 13, 153–166.
- Stevens, S. S., & Harris, J. R. (1962). The scaling of subjective roughness and smoothness. Journal of Experimental Psychology, 64, 489–494.
- Sur, M., Garraghty, P. E., & Bruce, C. J. (1985). Somatosensory cortex in macaque monkeys: Laminar differences in receptive field size in areas 3b and 1. Brain Research, 342, 391–395.
- Sur, M., Merzenich, M. M., & Kaas, J. H. (1980). Magnification, receptive-field area, and hypercolumn size in areas 3b and 1 of somatosensory cortex in owl monkeys. Journal of Neurophysiology, 44, 295–311.
- Sur, M., Wall, J. T., & Kaas, J. H. (1984). Modular distribution of neurons with slowly adapting and rapidly adapting responses in area 3b of somatosensory cortex in monkeys. Journal of Neurophysiology, 51, 724–744.
- Taylor, M. M., & Lederman, S. J. (1975). Tactile roughness of grooved surfaces: A model and the effect of friction. Perception and Psychophysics, 17, 23–36.
- Van Boven, R. W., & Johnson, K. O. (1994). The limit of tactile spatial resolution in humans: Grating orientation discrimination at the lip, tongue and finger. Neurology, 44, 2361–2366.
- Vega-Bermudez, F., & Johnson, K. O. (1999a). SA1 and RA receptive fields, response variability, and population responses mapped with a probe array. Journal of Neurophysiology, 81, 2701–2710.
- Vega-Bermudez, F., & Johnson, K. O. (1999b). Surround suppression in the responses of primate SA1 and RA mechanoreceptive afferents mapped with a probe array. Journal of Neurophysiology, 81, 2711–2719.
- Vega-Bermudez, F., & Johnson, K. O. (2001). Differences in spatial acuity between digits. Neurology, 56, 1389–1391.
- Vega-Bermudez, F., Johnson, K. O., & Hsiao, S. S. (1991). Human tactile pattern recognition: Active versus passive touch, velocity effects, and patterns of confusion. Journal of Neurophysiology, 65, 531–546.
- Warren, S., Hämäläinen, H. A., & Gardner, E. P. (1986). Objective classification of motion- and direction-sensitive neurons in primary somatosensory cortex of awake monkeys. Journal of Neurophysiology, 56, 598–622.
- Weisenberger, J. M. (1986). Sensitivity to amplitude-modulated vibrotactile signals. Journal of the Acoustical Society of America, 80, 1707–1715.
- Whang, K. C., Burton, H., & Shulman, G. L. (1991). Selective attention in vibrotactile tasks: Detecting the presence and absence of amplitude change. Perception and Psychophysics, 50, 157–165.
- Whitsel, B. L., Petrucelli, L. M., & Werner, G. (1969). Symmetry and connectivity in the map of the body surface in somatosensory area II of primates. Journal of Neurophysiology, 32, 170–183.
- Wohlert, A. B. (1996). Tactile perception of spatial stimuli on the lip surface by young and older adults. Journal of Speech and Hearing Research, 39, 1191–1198.
- Woolsey, C. N. (1943). ‘Second’ somatic receiving areas in the cerebral cortex of cat, dog, and monkey. Federation Proceedings, 2,
- Yoshioka, T., Dorsch, A. K., Hsiao, S. S., & Johnson, K. O. (2001). Neural coding mechanisms underlying perceived roughness of finely textured surfaces. Journal of Neuroscience, 21, 6905– 6916.
- Zhang, H. Q., Murray, G. M., Turman, A. B., Mackie, P. D., Coleman, G. T., & Rowe, M. J. (1996). Parallel processing in cerebral cortex of the marmoset monkey: Effect of reversible SI inactivation on tactile responses in SII. Journal of Neurophysiology, 76, 3633–3655.
- Zhou, Y. D., & Fuster, J. M. (1996). Mnemonic neuronal activity in somatosensory cortex. Proceedings of the National Academy of Sciences, USA, 93, 10533–10537.
- Zhou, Y. D., & Fuster, J. M. (1997). Neuronal activity of somatosensory cortex in a cross-modal (visuo-haptic) memory task. Experimental Brain Research, 116, 551–555.
- Zompa, I. C., & Chapman, C. E. (1995). Effects of cross-modal manipulations of attention on the ability of human subjects to discriminate changes in texture. Somatosensory and Motor Research, 12, 87–102.
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality