
View sample Neural Basis of Working Memory Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom writing service for professional assistance. We offer high-quality assignments for reasonable rates.
In experimental psychology, the term ‘working memory’ has been used to refer to the temporary storage of information that is necessary for cognitive processing underlying the comprehension and learning of new material, reasoning, mental arithmetic, etc. This theoretical construct emerged in the 1970s out of earlier experimental behavioral work on short-term memory (see Baddeley 1986 for a review). The term working memory has been defined variously to refer not only to the transient maintenance of information but also to its processing. For instance, one of the most popular working memory models in cognitive psychology assumes the existence of a central executive system supplemented by ‘slave’ systems that are specialized for the maintenance and processing of verbal and visual material (see Baddeley 1986). Such an expanded concept of a working memory system becomes tantamount to the entire neural processing of information that occupies current awareness. Naturally, any system defined so broadly will be the result of processing in multiple areas of the brain. This article will focus on research that aims to identify the role of several cortical structures in what has been termed working memory at the cognitive level of analysis.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. The Frontal Cortex And Working Memory
There is considerable evidence that the frontal cortex plays a major role in working memory although its precise role remains a matter of debate. The frontal cortex occupies a very large area of the primate cerebral cortex. Its posterior part, which lies immediately in front of the central sulcus, consists of the motor and premotor cortex (see Fig. 1). The vast area that lies further forward is often referred to as the ‘prefrontal’ cortex. The prefrontal cortex has expanded enormously in the primate brain and comprises many distinct areas. These areas differ in terms of their cytoarchitecture (i.e., the arrangement of the neurons within them) and have their own distinct connections with other parts of the brain (Pandya and Yeterian 1985). These anatomical facts imply that there must be some differences in the local neuronal computations occurring within particular areas since both the local circuitry and the inputs and outputs are distinct.
The first evidence that suggested that the prefrontal cortex plays a major role in working memory came from work in the 1930s by Jacobsen and co-workers who examined the performance of monkeys with lesions restricted to the prefrontal cortex on the delayed response task (Jacobsen 1936). This work showed that frontal cortical damage rendered animals incapable of performing the delayed response task which was introduced by Hunter in 1913 to study transient memory in animals. This task typically is conducted in the following manner. The experimenter attracts the monkey’s attention and, in full view of the animal, a reward (i.e., a piece of food) is placed under one of two identical stimuli (e.g., cups, plaques, etc.). The stimuli are then hidden from the animal’s view by an opaque screen and, after a delay of a few seconds, the screen is raised and the animal is allowed to choose one of the two stimuli to obtain the reward if, of course, the choice is correct. In interpreting the impairment on delayed response by monkeys with frontal cortical damage, Jacobsen (1936, p. 52) argued that the behavior of such animals ‘instead of being directed by a balance of immediate, recent and temporally remote stimulation, is dominated by present external stimuli and the stable habit systems derived from earlier training; the mnemonic influence of past experience is lacking.’
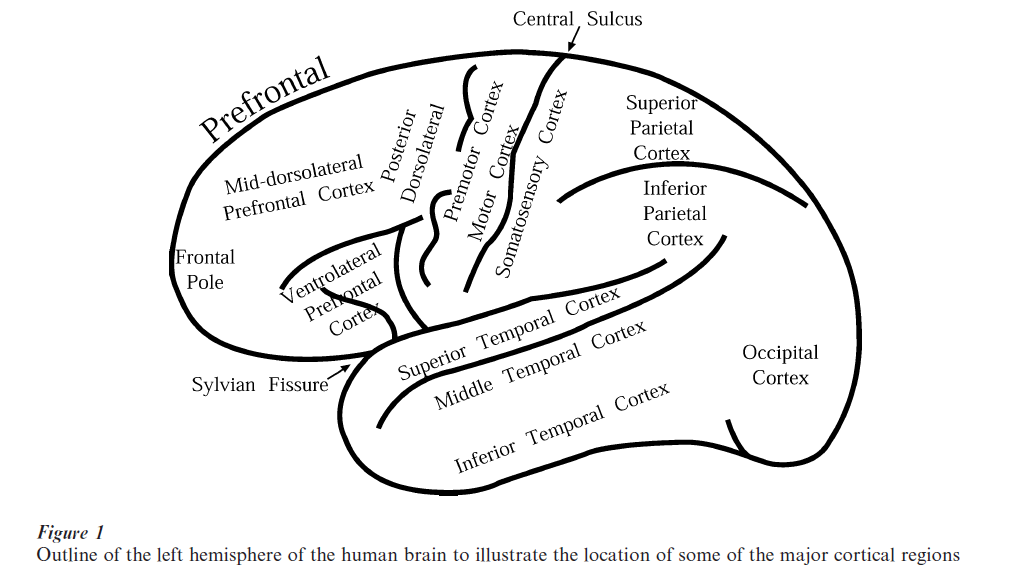
The demonstration that adequate performance of the delayed response and related tasks is severely impaired by lesions of the prefrontal cortex has been the starting point of a large amount of experimental work on its possible role in working memory. The lesion investigated in Jacobsen’s work involved the entire prefrontal cortex. Subsequent work in the monkey by Pribram, Mishkin, Gross, Weiskrantz, and others showed that the lateral surface of the prefrontal cortex was critical for delayed response performance. Furthermore, the deficit on the spatial version of the task was localized around the sulcus principalis (a prominent sulcus that runs longitudinally in the middle of the lateral frontal cortex of the monkey). It also became clear that the delay was a critical variable for the impairment on these tasks (see Petrides 1994 for a review).
By the early 1970s, the method of recording the activity of single neurons in alert monkeys as they performed various cognitive tasks was perfected and a number of studies appeared that recorded the activity of single neurons in the prefrontal cortex during delayed response performance. Fuster and Alexander (1971) in the United States and Kubota and Niki (1971) in Japan were the first to record such neuronal activity. This work demonstrated that certain neurons in the lateral prefrontal cortex respond specifically when a particular location is indicated as the one to be remembered and continue to discharge during the delay period until the choice has been made. Thus, these neuronal responses appear to maintain temporarily the processed information. This early work and several later studies by Goldman-Rakic, Miller, and co-workers clearly established the existence within the prefrontal cortex of sustained neuronal activity during the delay period of various tasks (see Fuster 1997 for a review).
Thus, both the lesion and the single-neuron work indicated the involvement of the prefrontal cortex in some aspect of working memory. But what exactly is the role of the prefrontal cortex in working memory? One of the theoretical interpretations of the above data was to assume that the prefrontal cortex is a critical region for the short-term maintenance of information and to conceive the different prefrontal cortical areas as specialized working memory modules for the maintenance of different types of information (Goldman-Rakic 1996). For instance, it was argued that the dorsolateral prefrontal area is specialized for spatial working memory (i.e., the maintenance of spatial information), whereas the ventrolateral prefrontal area is a specialized module for nonspatial visual working memory (Goldman-Rakic 1996).
An alternative theoretical model of the role of the different lateral prefrontal cortical areas in working memory was proposed by Petrides (1994), based on work that further analyzed the nature of the impairments on memory tasks by human patients and monkeys with prefrontal lesions. It was argued that the existence of delay neurons in the prefrontal cortex, although implying that some neuronal computations are occurring there during the delay, does not require the conclusion that the primary role of the prefrontal cortex is the short-term maintenance of information. For instance, neurons with sustained activity during the delay in memory tasks can be observed in virtually every part of the cerebral cortex, as well as in several subcortical structures. In addition, there is the major puzzle that patients with damage to the lateral prefrontal cortical region can perform well on several standard short-term memory tests, such as simple recognition, digit span, and story recall. These patients, however, are impaired on the self-ordered working memory tasks in which successful performance cannot be based simply on the maintenance of information in short-term memory but requires, in addition, the executive process of monitoring that information. In the self-ordered working memory tasks, the subjects are presented with different arrangements of the same set of stimuli and, on each trial, they have to select a different stimulus until all have been selected once. Successful performance requires the subjects to compare carefully those stimuli that they have already selected with those still remaining to be selected. In other words, events in working memory must be monitored closely. It was, therefore, argued that the problem of patients with prefrontal lesions lies not in memory per se, but in the application of executive processes on certain memory tasks (Petrides 1994, 2000).
In work on the monkey, where lesions can be restricted with precision within particular regions of the brain, it was demonstrated that damage to the mid- dorsolateral part of the prefrontal cortex results in severe impairments on these self-ordered tasks and related tasks that require the monitoring of externally ordered stimuli (see Petrides 1994). Further evidence of the importance of the mid-dorsolateral prefrontal cortex to the monitoring of information within working memory comes from functional neuroimaging studies on normal human subjects that have consistently shown increased activity in the mid- dorsolateral prefrontal region when the monitoring requirements of a memory task are high (Petrides 2000). In a study by Courtney et al. (1998) in which monitoring and manipulation were minimized, activity related specifically to the maintenance of an active mnemonic representation of spatial information was observed in a region that was posterior to the mid- dorsolateral prefrontal region, suggesting that posterior dorsolateral frontal cortex may play a role in the focussing of attention on information that is held in posterior cortical regions.
2. The Temporal And Parietal Cortex And Working Memory
A large number of studies with patients who had sustained lesions limited to posterior cortical regions have documented impairments in short-term memory, thus providing evidence that the temporary maintenance of information may be occurring in posterior cortical regions (see Vallar and Shallice 1990). There are several reports of patients with a selective impairment in auditory–verbal short-term memory after lesions in the inferior parietal region of the left hemisphere. These patients may be able to maintain and repeat single numbers, letters, or words presented auditorily, but their span (i.e., the number of items that they can maintain after a single presentation) is severely limited (e.g., Warrington and Shallice 1969). Patients with severely impaired short-term memory spans for visual verbal material usually have lesions in the occipitotemporal region of the left hemisphere (e.g., Shallice and Saffran 1986). De Renzi et al. (1977) have investigated visuo-spatial span using the Corsi blocks test. In this test, the patient observes the examiner tap, in sequence, a number of randomly arranged blocks and has to reproduce this sequence. Patients with right posterior hemisphere lesions were particularly impaired on this task, suggesting that the occipitoparietal cortex of the right hemisphere may be a key region for the maintenance of visuo-spatial information (De Renzi et al. 1997).
This notion that posterior cortical regions may play a role in the maintenance of information in working memory receives additional support from work in the monkey. For instance, there is considerable evidence that the anterior part of the inferior temporal cortical region, which is known to be the final stage in the occipitotemporal neural system underlying visual object perception, is also critically involved in shortterm visual memory. This evidence comes both from investigations of the effects of lesions (Mishkin 1982) and from studies in which the activity of single neurons was recorded in monkeys as they performed visual short-term memory tasks (Fuster and Jervey 1982, Miller et al. 1991). Similar evidence exists in other sensory modalities. Lesions of certain parts of the superior temporal cortex have been shown to lead to a serious impairment in auditory short-term memory (e.g., Colombo et al. 1990).
How can the evidence for impairments in working memory both from posterior cortical regions and prefrontal cortex be conceptualized? In one theoretical model, the temporary storage and processing of incoming and recalled information is seen as an integral part of neuronal computation in modality specific and multimodal posterior association cortical areas (Petrides l994). In other words, neural activity in these posterior cortical areas underlies not only perceptual processing and long-term storage of information, but also short-term memory of recently processed information which may be currently provided by the environment or re-activated from longterm storage. The prefrontal cortical areas are considered to play a role in working memory as distinct executive processing modules that allocate attention to different types of information, and monitor the contents of information in posterior cortical regions. Thus, if working memory were to be viewed as the multicomponent workspace of current cognitive activity, then this workspace can be construed as the local computations occurring within specialized posterior cortical regions for the maintenance and processing of modality-specific information, within frontal cortical regions for different aspects of executive processing and the bi-directional interactions between them.
Bibliography:
- Baddeley A D 1986 Working Memory. Oxford University Press, Oxford, UK
- Colombo M, D’Amato M R, Rodman H R, Gross C G 1990 Auditory association cortex lesions impair auditory short-term memory in monkeys. Science 247: 336–8
- Courtney S M, Petit L, Maisog J Ma, Ungerleider L G, Haxby J V 1998 An area specialized for spatial working memory in human frontal Science 279: 1347–51
- De Renzi E, Faglioni P, Previdi P 1977 Spatial memory and hemispheric locus of lesion. Cortex 13: 424–33
- Fuster J M 1997 The Prefrontal Anatomy, Physiology and Neuropsychology of the Frontal Lobe. Raven Press, New York
- Fuster J M, Alexander G E 1971 Neuron activity related to short-term memory. Science 173: 652–4
- Fuster J M, Jervey J P 1982 Neuronal firing in the inferotemporal cortex of the monkey in a visual memory Journal of Neuroscience 2: 361–75
- Goldman-Rakic P S 1996 Architecture of the prefrontal cortex and the central executive. Annals of the New York Academy of Science 769: 71–83
- Jacobsen C F 1936 Studies of cerebral function in I. The functions of the frontal association areas in monkeys. Comparative Psychology Monographs 13: 1–60
- Kubota K, Niki H 1971 Prefrontal cortical unit activity and delayed alternation performance in monkeys. Journal of Neurophysiology 34: 337–47
- Miller E K, Li L, Desimone R 1991 A neural mechanism for working and recognition memory in inferior temporal Science 254: 1377–9
- Mishkin M 1982 A memory system in the monkey. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences 298: 85–95
- Pandya D N, Yeterian E H 1985 Architecture and connections of cortical association In: Peters A, Jones E G (eds.) Cerebral Cortex. Plenum Press, New York
- Petrides M 1994 Frontal lobes and working memory: Evidence from investigations of the effects of cortical excisions in nonhuman primates. In: Boller F, Grafman J (eds.) Handbook of Neuropsychology. Elsevier, Amsterdam, Vol. 9, pp. 59–82
- Petrides M 2000 Frontal lobes and In: Boller F, Grafman J (eds.) Handbook of Neuropsychology, 2nd edn. Elsevier, Amsterdam, Vol. 2, pp. 67–84
- Shallice T, Saffran E M 1986 Lexical processing in the absence of explicit word identification: Evidence from a letter-by-letter reader. Cognitive Neuropsychology 3: 429–58
- Vallar G, Shallice T (eds.) 1990 Neuropsychological Impairments of Short-term Memory. Cambridge University Press, Cambridge, UK
- Warrington E K, Shallice T 1969 The selective impairment of auditory verbal short-term memory. Brain 92: 885–96
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality