
View sample Neural Basis Of Syntactic Aspects Research Paper. Browse other research paper examples and check the list of research paper topics for more inspiration. If you need a religion research paper written according to all the academic standards, you can always turn to our experienced writers for help. This is how your paper can get an A! Feel free to contact our custom writing service for professional assistance. We offer high-quality assignments for reasonable rates.
Natural languages are built on three different knowledge components: the sound of words, called phonology; the meaning of words, called semantics; and the grammatical rules according to which words are put together, called syntax. This research paper concerns the neural mechanism underlying the knowledge and use of syntax. Three methods have been used to study the neural basis of syntax: (a) the observation of the correlation between brain lesion and behavior; (b) functional imaging approaches; and (c) event-related brain potential methods. Evidence from these three approaches with respect to syntactic processing will be reviewed and an explanatory model covering major aspects of the data will be suggested.
Academic Writing, Editing, Proofreading, And Problem Solving Services
Get 10% OFF with 24START discount code
1. Lesion–Behavior Approaches
Lesion–behavior approaches to investigating the neural basis of language date back to the middle of the nineteenth century (Broca 1861, Wernicke 1874). Using such an approach it was observed that patients with lesions in the temporal region of the left hemisphere (the so-called Wernicke’s area, Brodmann’s area, BA 22/42) are characterized by poor comprehension, whereas their speech is fluent and often syntactically well structured although semantically empty. Patients with lesions in the left inferior frontal gyrus (the so-called Broca’s area, BA 44), in contrast, demonstrate a selective deficit for syntactic aspects in language production and in language comprehension. Patients with lesions in Broca’s area produce agrammatic speech, that is utterances containing content words, but lacking most of the grammatical morphology (function words and inflections). They fail to comprehend sentences whenever the correct interpretation depends on the full and adequate processing of the grammatical morphology. In the 1980s a central topic of discussion centered around three possible ways of understanding this behavior. A first view took this behavior to be due to a lack of grammatical knowledge as such (Caramazza and Zurif 1976, Berndt and Caramazza 1980). A second approach interpreted it as being caused by computational deficit either in accessing lexical information or in accessing the syntactic information encoded in function words and inflectional morphology (Bradley et al. 1980). A third view assumed the computational deficit to be based either on reduced memory resources (Kolk and Van Grunsven 1985) or on reduced processing resources leading to a trade-off between syntactic and semantic interpretative processes (Linebarger et al. 1983).
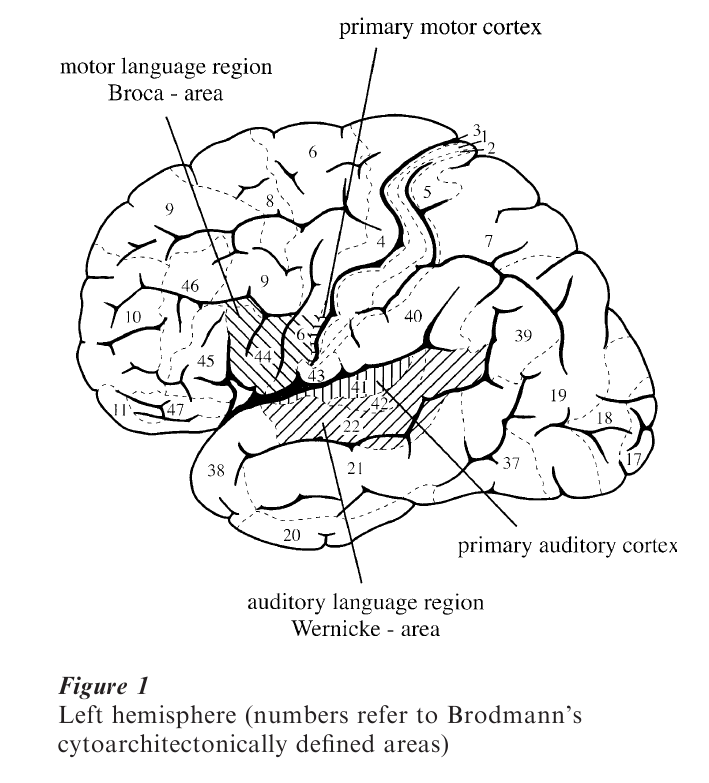
It has been shown that Broca’s patients are able to judge a sentence’s grammaticality (Linebarger et al. 1983) and even show grammatical priming (i.e., grammatically correct context sentences enhanced the recognition of a target word compared to ungrammatical context sentences; see Haarmann and Kolk 1991). These findings lead to the assumption that Broca’s area may not be the brain area where syntactic knowledge is represented, but rather that this brain tissue supports syntactic computation. While some view this brain region to be the home of very specific syntactic computations necessary to process dislocated elements (e.g., moved constituents that leave behind traces, as Grodzinsky 2000), as others consider this brain area to support on-line processes of syntactic phrase-structure building (Friederici 1995).
There is, however, evidence that might challenge these views. It has been reported that patients with lesions outside Broca’s area show syntactic comprehension deficits in off-line tasks (Caplan et al. 1985, 1996). Moreover, both Broca’s and Wernicke’s patients display a certain similarity in their perfomance in offline comprehension tasks (Bates et al. 1987). It was demonstrated that patients speaking a language which marks syntax by inflectional morphology are generally more deficient, irrespective of their classification, than those speaking a language marking syntax by strict word order. However, the finding that in a highly inflected language Broca’s patients are more affected than Wernicke’s patients suggests a particular deficit of Broca’s aphasics for syntactic morphology. Thus it appears that similarities between different patient groups can be observed when using off-line comprehension tasks. As these tasks only reflect the end point of the comprehension process they may imply certain on-line operations possibly supported by Broca’s area or adjacent tissue, as well as other operations possibly supported by other areas (see also Zurif et al. 1993) (see Fig. 1).
2. Functional Neuroimaging
New imaging techniques during the 1990s have provided additional insight into the neural basis of syntactic parsing. Positron emission tomography (PET) studies (see Sect. 4.3.3) have investigated the reading of syntactically simple and complex sentences. It was found that the regional blood flow increases in the left Broca’s area as a function of syntactic complexity (Stromswold et al. 1996). A recent study using functional magnetic resonance imaging (f MRI) to register the brain’s activity during the reading of sentences of different complexity found a significant increase of the hemodynamic response in the left Broca’s and Wernicke’s area as a function of the sentence’s complexity (Just et al. 1996).
Other studies investigated the brain–syntax relationship by comparing the brain activity during the processing of word lists with that during sentence processing. In an event-related fMRI study Friederici et al. (2000) presented four different auditory stimulus types: a list of real words (nouns); a list of pseudowords; normal sentences containing function words and real content words; and sentences containing function words and pseudowords instead of real content words. Both sentence types differed from the two word lists, showing an additional activation in the anterior portion of the left superior temporal gyrus. The sentences that contained only grammatical morphemes and pseudowords instead of content words displayed an additional activation in the left frontal operculum in the vicinity of the Broca’s area (i.e., medial to BA 44).
The combined imaging data suggest that parts of the left superior temporal gyrus and the left frontal operculum are the brain systems that support syntactic processes. When speculating about the specific contribution of the two brain regions one may again consult the patient data. Given the observation that Broca’s patients are able to perform grammaticality judgments (Linebarger et al. 1983), but are deficient in computing syntactic information on-line (Friederici 1995), and the finding that Wernicke’s patients are deficient in performing judgments (Blumstein et al. 1982), but perform similar to normals in some on-line comprehension tasks (Zurif et al. 1993), the following account may be formulated. While the left temporal region is necessary for the representation of syntactic knowledge, the left inferior frontal region supports syntactic procedures in real time.
3. Neurophysiological Approaches
The temporal course of the neural activity underlying syntax processing has been evaluated by means of electroencephalography (EEG) (see Sects. 4.3.5 and 4.3.6) and more recently also by means of magnetoencephalography (MEG) (see Sect. 4.3.7). With their seminal paper Kutas and Hillyard (1983) introduced the event-related brain potential (ERP) method to investigate language processes. They observed a particular ERP component correlated with lexicalsemantic processes: a negative potential peaking around 400 msec after the onset of the critical event, called the N400 component. The critical event in their study was a sentence final word which was highly unexpected and semantically incoherent (for example: ‘He spread his bread with butter and socks.’). It was shown that the amplitude of the N400 varied as a function of a word’s semantic expectancy and also as a function of a word’s lexical frequency. Importantly, the N400 component has been reported in a number of different languages including English, German, Dutch, French, Hebrew, and even sign language in the visual as well as in the auditory domain (for a review, see Van Petten 1995). Note that unexpected events in a non-semantic domain elicit a positivity (P300) rather than an N400 (Donchin 1981).
3.1 Syntax-Related Electrophysiological Correlates
The picture is somewhat more complex for the ERP pattern correlated with syntactic processing. There are two syntax-related components that have been identified: an early left anterior negativity, and a late centroparietal positivity. While the anterior negativity is only present when processing outright violations, the late positivity is observed when processing outright violation and when processing violations of structural preferences.
3.1.1 The Syntax-Related Late Positivity. The late positivity is a centroparietally distributed positivity usually at around 600 msec and beyond, called P600. P600 components are observed both in correlation with outright syntactic violation and with violations of syntactic preferences. Violations of the latter kind are present in so-called ‘garden path’ sentences, that is sentences for which it becomes clear at a particular word that the perceiver was misguided structurally by following a simple instead of a more complex syntactic structure (for example: ‘The broker hoped to sell the stock was sent to jail’ vs. ‘The broker persuaded to sell the stock was sent to jail,’ Osterhout and Holcomb 1992). At critical points in the sentence (at the word ‘to’ and the word ‘was’) the reader listener has to revise and reanalyze the structure initially followed. The P600 is also present in sentences containing an outright syntactic violation, be it a phrase-structure violation realized by a word category error (Neville et al. 1991, Friederici et al. 1993) or be it a morphosyntactic violation realized by an incorrect inflection (Gunter et al. 1997, Munte et al. 1997). Functionally, the P600 is taken to reflect costs of reprocessing (Osterhout et al. 1994), syntactically guided processes of reanalysis and repair (Friederici 1995), or syntactic integration difficulties in general (Kaan et al. 2000).
3.1.2 The Syntax-Related Anterior Negativity. For outright syntactic violations an anterior negativity, that is, a frontally distributed negative waveform usually with a maximum over the left, is observed. It is called ELAN (early left anterior negativity) or LAN (left anterior negativity), depending on its latency of occurrence. Before we discuss the particular appearance of this syntax-related negativity in more detail we shall recapitulate briefly that syntactic information is multiply encoded in a given language.
3.2 Domains Of Syntax And Their Neurophysiological Correlates
There are at least four syntactic domains that have to be considered: (a) word category information necessary to build up local syntactic structures; (b) morphosyntactic information such as verb inflections necessary to establish agreement between different elements within and across phrases or case markings allowing the assignment of thematic roles (who is doing what to whom); (c) verb argument structure information necessary to establish subcategorization relations; and (d) long-distance dependencies necessary to establish the relation between a moved element (filler: ‘who’) and its original position in the sentence (gap: ) in a sentence such as ‘Who did Peter see ?’ Some, but not all, aspects of these different domains have been investigated using neurophysiological recordings. We shall take up the different domains in turn.
3.2.1 Word Category Information. The anterior negativity was found in correlation with phrase-structure violations realized as word category errors and with the processing of function words. It was observed between 125 and 200 msec after the critical word’s onset (italicized) in a number of studies using auditory presentation (for example: ‘Der Freund wurde besucht’/‘The friend was in the visited’ vs. for ‘Der Freund wurde besucht’ ‘The friend was visited,’ Friederici et al. 1993, Hahne and Friederici 1999). (But see Neville et al. 1991, who found an early and late negativity in the visual domain.) The distribution of the word category error related negativity is often frontal with a maximum to the left, but has also been found to be less lateralized (Friederici et al. 1999). So far it is not yet clear what determines the distributional differences. Current research is trying to solve this issue. With respect to the possible neural generators for the anterior negativity it is interesting that left anterior cortical lesions eliminate the anterior word category related negativity whereas left anterior lesions restricted to the basal ganglia do not (Friederici et al. 1999). Functionally, the word category related negativity was interpreted as reflecting initial structure building processes (Friederici 1995), which according to syntax first models (Frazier 1987), are based on word category information only.
A comparable left anterior negativity also related to word category information was observed for the processing of function words in syntactically correct sentences (Neville et al. 1992, see also Pulvermuller et al. 1995). It was present around 280 msec post onset when reading sentences. Similar to the above discussed negativity, the function word related negativity was eliminated with left anterior lesions (Ter Keurs et al. 1999). In line with the proposal by Friederici (1995), the authors interpret the left anterior negativity to reflect processes of word category identification.
3.2.2 Morphosyntactic Information. Morphosyntactic violations (for example: ‘The plane took we to paradise’ vs. ‘The plane took us to paradise’) usually elicit a left anterior negativity between 300 and 500 msec after the critical word’s onset (italicized). It was found in the visual domain in a number of different languages (Munte et al. 1993, Gunter et al. 1997, Coulson et al. 1998), as well as in the auditory domain (although with a somewhat larger distribution, Friederici et al. 1993). Note that the types of morphosyntactic violations investigated in these studies do not alter the critical word’s syntactic category, but influence the relation between the verb and its arguments (e.g., subject–verb agreement). Under the view that initial phrase-structure building is concerned primarily with word category information, and that the lexical information encoded in an inflection only becomes relevant when thematic roles are to be assigned, it is conceivable that the violation of the former information type elicits an earlier brain response than the violation of the latter information type.
3.2.3 Verb Argument Structure Information. The processing of verb argument structure information has been investigated in a number of different studies using different types of violation. In an early study, Rosler et al. (1993) registered ERP for sentences which contained subcategorization violations (for example: ‘Der Lehrer wurde gehustet’ ‘The teacher was coughed’). A left anterior negativity between 300 and 500 msec was found in correlation with the violation. In this study, the sentence final verb (one-place verb) does not match the thematic grid opened up by the passive construction (two-place verb). When the mismatch involved subject vs. object arguments, again a left anterior negativity was reported (Coulson et al. 1998). Thus it appears that violations concerning the relation between external (subject) and internal arguments (objects) also evoke a left anterior negativity.
Interestingly, the argument structure related negativity is present between 300 and 500 msec, that is, the time domain in which the lexical-semantic ERP component is found. This is not surprising, given that the availability of the verb argument structure information requires full access to the verb’s lexical entry.
3.2.4 Long-Distance Dependencies. Long-distance dependencies that establish the relation between a moved constituent (filler) and its original position (gap) have been investigated by Kluender and Kutas (1993) in a number of constructions. They found a LAN at the gap position at which the previously perceived filler has to be reactivated. As it is assumed that the filler has to be kept in memory until the gap is met, they claim that the LAN is a reflection of verbal memory. The two functional interpretations of the LAN as a syntactic violation related LAN and a filler-gap related LAN has recently been solved by showing that the former is present locally (at the violation) and that the latter spans from the filler to the gap position over the sentence (Kluender et al. 1998).
In conclusion, it appears that outright syntactic violations elicit a local anterior negativity with a maximum over the left hemisphere, in addition to a P600. Moreover, the combined data suggest that word category related anterior negativities are present earlier than anterior negativities found in correlation with lexically encoded morphosyntactic information and lexically encoded verb argument information.
4. Modeling The Neurocognition Of Syntactic Processes
The data provided by recent ERP and functional imaging studies have advanced our knowledge on the neural basis of language, and of syntax in particular. The network identified to support syntactic processes clearly includes superior temporal and inferior frontal areas dominantly in the left hemisphere. Their specific function still has to be determined in detail. It appears, however, that the left temporal region is relevant for the representation of syntactic knowledge, whereas the inferior frontal region supports syntactic procedures. The timing of the brain’s activity correlated with syntactic processes can be described to consist of different stages: an early stage during which first-pass parsing processes (based on word category information) take place, and a next stage during which lexically encoded syntactic information is processed. Both are reflected in a left anterior negativity in the ERP. A late stage of syntactic processes is reflected by a late positivity in the ERP. This component is observed whenever syntactic integration is difficult. Future research will have to determine whether different aspects of syntactic integration difficulties are correlated with distinct topographies of the late positivity which would indicate the involvement of different neural structures.
Bibliography:
- Bates E, Friederici A D, Wulfeck B 1987 Comprehension in aphasia: A cross-linguistic study. Brain and Language 32: 19–67
- Berndt R S, Caramazza A 1980 A redefinition of the syndrome of Broca’s aphasia: Implications for a neuropsychological model of language. Applied Psycholinguistics 1: 225–78
- Blumstein S E, Milberg W, Schrier R 1982 Semantic processing in aphasia: Evidence from an auditory lexical decision task. Brain and Language 17: 301–15
- Bradley D C, Garrett M F, Zurif E B 1980 Syntactic deficits in Broca’s aphasia. In: Caplan D (ed.) Biological Studies on Mental Processes. MIT Press, Cambridge, MA, pp. 269–86
- Broca P 1861 Remarques sur le siege de la faculte du langage articule, suivies d’une observation d’aphemie (perte de la parole). Bulletins de la Societe Anatomique de Paris 6: 330– 57
- Caplan D, Baker C, Dehaut F 1985 Syntactic determinants of sentence comprehension in aphasia. Cognition 21: 117–75
- Caplan D, Hildebrandt N, Makris N 1996 Location of lesions in stroke patients with deficits in syntactic processing in sentence comprehension. Brain 119: 933–49
- Caramazza A, Zurif E B 1976 Dissociation of algorithmic and heuristic processes in language comprehension: Evidence from aphasia. Brain and Language 3: 572–82
- Coulson S, King J, Kutas M 1998 Expect the unexpected: Event- related brain responses of morphosyntactic violations. Language and Cognitive Processes 13: 21–58
- Donchin E 1981 Surprise!…Surprise? Psychophysiology 18: 493–513
- Frazier L 1987 Theories of sentence processing. In: Garfield J (ed.) Modularity in Knowledge Representation and Naturallanguage Processing. MIT Press, Cambridge, MA
- Friederici A D 1995 The time course of syntactic activation during language processing: A model based on neuropsychological and neurophysiological data. Brain and Language 50: 259–81
- Friederici A D, Meyer M, Von Cramon D Y 2000 Auditory language comprehension: An event-related fMRI study on the processing of syntactic and lexical information. Brain and Language 74: 289–300
- Friederici A D, Pfeifer E, Hahne A 1993 Event-related brain potentials during natural speech processing: Effects of semantic, morphological and syntactic violations. Cognitive Brain Research 1: 183–92
- Friederici A D, Von Cramon D Y, Kotz S A 1999 Language related brain potentials in patients with cortical and sub-cortical left hemisphere lesions. Brain 122: 1033–47
- Grodzinsky Y 2000 The neurology of syntax: Language use without Broca’s area. Behavioral and Brain Sciences 23: 1–71
- Gunter T C, Stowe L A, Mulder G 1997 When syntax meets semantics. Psychophysiology 34: 660–76
- Haarmann H J, Kolk H H J 1991 Syntactic priming in Broca’s aphasics: Evidence for slow activation. Aphasiology 5: 247–63
- Hahne A, Friederici A D 1999 Electrophysiological evidence for two steps in syntactic analysis: Early automatic and late controlled processes. Journal of Cognitive Neuroscience 11: 193–204
- Just M A, Carpenter P A, Keller T A, Eddy W F, Thulborn K R 1996 Brain activation modulated by sentence comprehension. Science 274: 114–16
- Kaan E, Harris A, Gibson G, Holcomb P J 2000 The P600 as an index of syntactic integration diffi Language and Cogniti e Processes 15: 159–201
- Kluender R, Kutas M 1993 Subjacency as a processing phenomenon. Language and Cognitive Processes 8: 573–633
- Kluender R, Munte T, Wind-Cowles H, Szentkuti A, Walenski M, Wieringa B 1998 Brain potentials to English and German questions. Journal of Cognitive Neuroscience Supplement 24
- Kolk H H J, Van Grunsven M J F 1985 Agrammatism as a variable phenomen. Cognitive Neuropsychology 2: 347–84
- Kutas M, Hillyard S A 1983 Event-related potentials to grammatical errors and semantic anomalies. Memory & Cognition 11: 539–50
- Linebarger M C, Schwartz M F, Saffran E M 1983 Sensitivity to grammatical structure in so-called agrammatic aphasics. Cognition 13: 361–92
- Munte T F, Heinze H J, Mangun G R 1993 Dissociation of brain activity related to syntactic and semantic aspects of language. Journal of Cognitive Neuroscience 5: 335–44
- Munte T F, Matzke M, Johannes S 1997 Brain activity associated with syntactic incongruencies in words and pseudowords. Journal of Cognitive Neuroscience 9: 318–29
- Neville H J, Mills D L, Lawson D S 1992 Fractionating language: Different neural subsystems with different sensitive periods. Cerebral Cortex 2: 244–58
- Neville H J, Nicol J L, Barss A, Forster K I, Garrett M F 1991 Syntactically based sentence processing classes: Evidence from event-related brain potentials. Journal of Cognitive Neuroscience 3: 151–65
- Osterhout L, Holcomb P J 1992 Event-related potentials and syntactic anomaly. Journal of Memory and Language 31: 785–804
- Osterhout L, Holcomb P J, Swinney D 1994 Brain potentials elicited by garden-path sentences: Evidence of the application of verb information during parsing. Journal of Experimental Psychology-learning Memory and Cognition 20: 786–803
- Pulvermuller F, Lutzenberger W, Birbaumer N 1995 Electrocortical distinction of vocabulary types. Electroencephalography and Clinical Neurophysiology 94: 357–70
- Rosler F, Friederici A D, Putz P, Hahne A 1993 Event-related brain potentials while encountering semantic and syntactic constraint violations. Journal of Cognitive Neuroscience 5: 345–62
- Stromswold K, Caplan D, Alpert N, Rauch S 1996 Localization of syntactic comprehension by positron emission tomography. Brain and Language 52: 452–73
- Ter Keurs M, Brown C M, Hagoort P, Stegemann D F 1999 Electrophysiological manifestations of open-and closed-class words in patients with Broca’s aphasia with agrammatic comprehension: An event-related brain potential study. Brain 122: 839–854
- Van Petten C 1995 Words and sentences: Event-related brain potential measures. Psychophysiology 32: 511–25
- Wernicke C 1874 Der aphasische Symptomenkomplex. Cohn and Weigert, Breslau, Germany
- Zurif E B, Swinney D, Prather P, Solomon J, Bushell C 1993 An on-line analysis of syntactic processing in Broca’s and Wernicke’s aphasia. Brain and Language 45: 448–64
ORDER HIGH QUALITY CUSTOM PAPER

Always on-time

Plagiarism-Free

100% Confidentiality